
Crystallotesta marika, Hodgson, Chris J. & Łagowska, Bozena, 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.205749 |
|
DOI |
https://doi.org/10.5281/zenodo.6193120 |
|
persistent identifier |
https://treatment.plazi.org/id/03A487E9-FFB1-FFF9-E2CE-2EDF78334FAA |
|
treatment provided by |
Plazi |
|
scientific name |
Crystallotesta marika |
| status |
sp. nov. |
Crystallotesta marika sp. n.
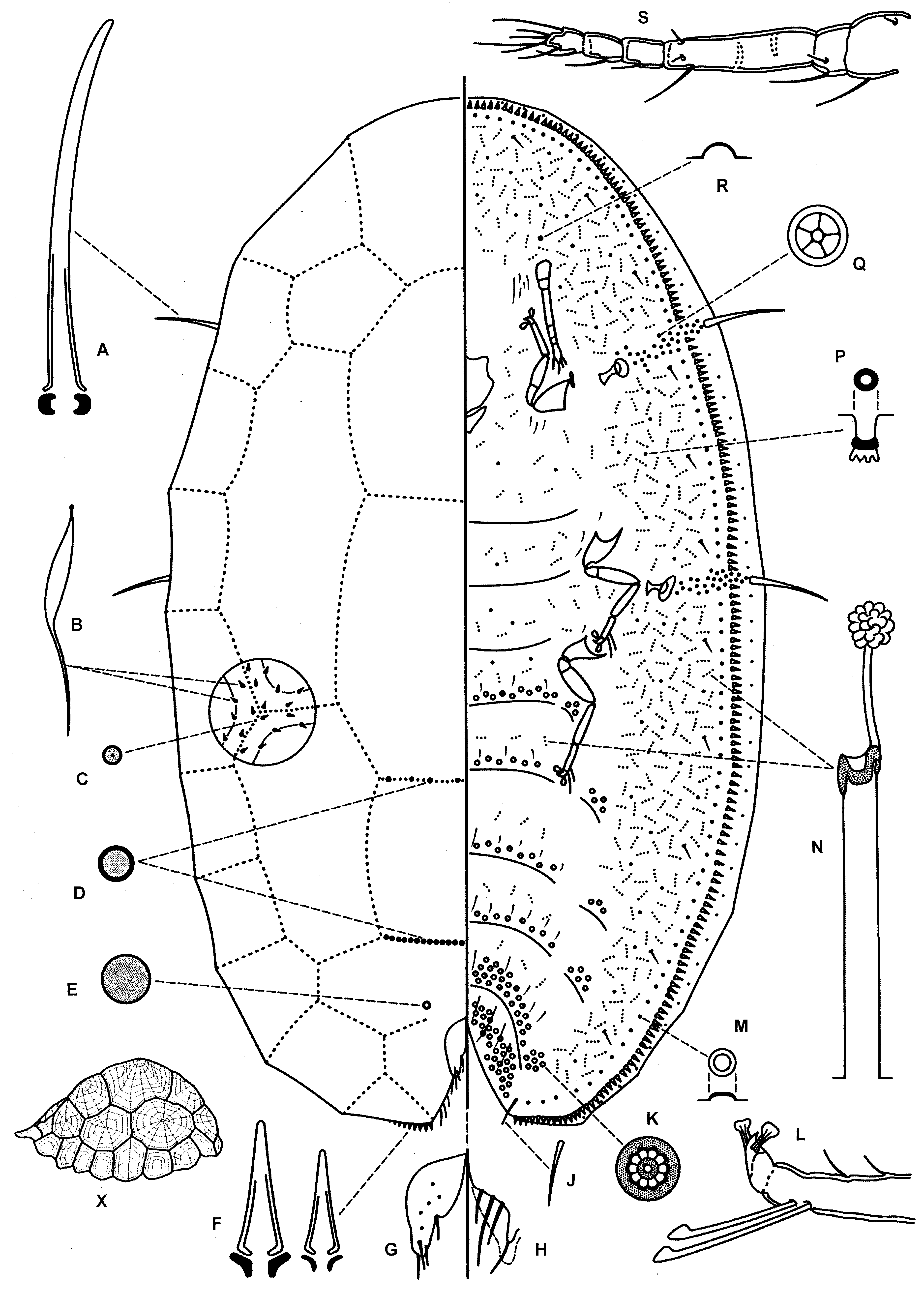
( Fig. 3 View FIGURE 3 )
Material examined. Holotype female: Fiji, Suva, Thurston Gardens, on epiphytic fern Pyrrosia adnascens (Swartz) Ching (Polypodiaceae) , 25.vii.2009, C.J. Hodgson ( BMNH): 1/1adf (good).
Paratype females: Fiji, as previous ( BMNH): 1/1adf (fair) + 1 pupa + parasite; as previous but collected 11.vii.2009 ( BMNH): 2/2 adff (poor) + 1 pupa + cast adm exuvia; and as previous but collected on 12.vii.2009 ( BMNH; USNM): 2/4adff (fair–good).
Adult female
Unmounted material. Test of adult female white, glassy, more or less oval, convex, quite narrow; formed of a series of polygonal wax plates, dorsal-most wax plate largest, marginal fringe of wax plates smallest; each plate marked with striae radiating from its apex. Length of test 2.4 mm, height 1.2 mm.
Mounted material. Body elongate-oval, equally rounded at both ends. Length of young adult female 2.0– 2.5 mm (older female – 5.3 mm); breadth of young adult female 1.1–1.4 mm (older female – 2.7 mm).
Dorsum. Derm membranous. Dorsal setae absent. With a series of 5 longitudinal rows of polygonal reticulation areas delineated by reticulation lines of pores; with 10 pairs of reticulation areas along lateral margins plus a large reticulation area anteriorly on head (i.e. with 21 marginal reticulation areas in all); also, on each side, with 4 reticulation areas mediolaterally, two of them large plus 3 large reticulation areas medially on dorsum; also with another small plate present just anterior to anal plates. Each reticulation line broad, composed of 5 parallel lines of pores; a median line with numerous very small, simple pores, each close together; plus, on each side, two lines of microductules, one close to median simple pore line, other some distance away, microductules much less frequent than simple pores. Dorsal pores of 4 types: (i) minute, dark microductules, each with a long inner ductule, present within outer 4 lines of pores in each reticulation line; (ii) very small simple pores of perhaps two sizes, smaller (about 1.5 µm wide) abundant in centre of each reticulation line; larger (about 2 µm wide) forming a sparse marginal line very close to marginal setae; frequency variable; (iii) larger, simple pores, each about 3 µm wide, present in two transverse reticulation lines just anterior to anal plates; and (iv) single, larger simple pores laterad to anterior margin of each anal plate, each about 4 µm wide. Preopecular pores, dorsal macropores, dorsal tubercles and dorsal tubular ducts absent. Anal plates each triangular with anterior margin rather shorter than posterior margin and with a rather pointed apex: each plate 110–135 µm long, combined widths 105–140 µm; with 3–6 minute pores on dorsal surface of each plate; each plate with 4 stoutly spinose setae, with 2 inner margin setae, each 13–25 µm, 1 apical seta 34–40 µm long and 1 dorsal seta 18 µm. Anogenital fold with 3 pairs on anterior margin setae plus 1 pair on lateral margins; longest 40–80 µm. Anal ring with 4 pairs of setae, each about 240–290 µm long.
Margin. Marginal setae strongly spinose, tapering to a sharp point and with a shallow basal socket, each 10–20 µm long; abundant, with 61–74 laterally between stigmatic clefts, those posterior to each cleft sometimes in a group 2 or 3 wide; also with 2 or 3 longer spinose setae along dorsal margin of each anal cleft; reticulation setae not noticeably differentiated. Each cleft with 1 long stigmatic spine, slightly tapering and curved, with a well-developed basal socket; length 113–138 µm. Eyespot not detected.
Venter. Membranous. Pregenital disc-pores with mainly 10 (6–10) loculi; number across each segment: (medially across segment/laterally on one side): VII, 19–26/16–19; VI, 47–51/6–7; V, 13–16/3–5; IV, 10–16/2–3; III, 9– 11/2–4; II, 13–17/1–3; sometimes also with 1–4 disc-pores between metacoxae. Spiracular disc-pores each with 5 loculi, in bands 1–5 pores wide; with 26–44 in each anterior band and 30–49 in each posterior band, each band extending onto dorsum to near each stigmatic spine; with none extending medially past peritreme. Ventral microducts of 1 type randomly distributed throughout. With 0 or 1 preantennal pore. Also with a line of simple pores around submargin, with 17–30 on each side between stigmatic clefts. Ventral tubular ducts of 1 type; each usually with a long outer ductule, long inner ductule and a well-developed terminal gland; present throughout but most abundant in a broad submarginal band. Ventral setae: each anal lobe with 1 rather spinose anal lobe seta 17–25 µm long; with 1 pair of anterior anal cleft setae; with long pregenital setae medially on pregenital segments VII, VI, and V, longest 65–68 µm long; number of setae medially on each abdominal segment: VII 8 long setae, no short setae; VI 8–10 setae (2–4 long setae + 6–8 moderately long setae); V 9–12 (2 long setae + 4–6 moderately setae + 1–6 short setae); IV 9–12 setae (2 moderately long setae + 9–10 short setae); III & II 3–6 short setae; also with 0 or 1 seta near each metacoxa and 1–3 setae near each meso- and procoxa; length of setae associated with each procoxa 10 µm; with 1–6 submarginal setae on each side between stigmatic areas; and with 3–5 pairs of interantennal setae.
Antennae each 6 segmented, with 1 or 2 pseudosegments in long third segment; total length 230–295 µm; scape with only 2 setae; length of apical seta 27–30 µm. Length of clypeolabral shield 138–168 µm. Width of spiracular peritremes: anterior 48–70 µm, posterior 53–75 µm. Legs without a tibio-tarsal articulatory sclerosis; tarsal digitules longer than claw digitules; claw without a denticle; lengths (metathoracic): coxa 125–147 µm, trochanter+femur 155–178 µm, tibia 117–130 µm, tarsus 65–85 µm, claw 13–18 µm. Vulva present in abdominal segment VII.
Comment. Crystallotesta marika fits extremely well into the genus Crystallotesta Henderson & Hodgson , which was thought to be restricted to New Zealand. In having long stigmatic spines, it is close to C. ornata (Maskell) and (particularly) C. ornatella Henderson & Hodgson but differs from both in having the following combination of characters: (i) 21 distinct reticulation areas around the margin; (ii) dorsal pores in each reticulation line mainly small simple pores, with larger small simple pores restricted to posterior 2 transverse reticulation lines; (iii) complete absence of dorsal macropores, and (iv) complete absence of dorsal tubular ducts.
In the key to Crystallotesta species in Hodgson and Henderson (2000), couplet 3 can be modified as follows to separate C. marika from C. ornata and C. ornatella :
3. Dorsal macropores either tubular or absent; with 2 or 3 pairs of setae along each margin of anal cleft; antennae 6-segmented................................................................................................... 3a. - Dorsal macropores present and highly convex; with only 2 pairs of setae on each anal cleft margin; antennae generally 8 segmented............................................................................... C. ornata (Maskell) 3a. Dorsal macropores tubular, frequent along most reticulation lines; with 7–10 simple pores in each ventral submarginal line between lateral stigmatic areas............................................... C. ornatella Henderson & Hodgson - Dorsal macropores absent; with 17–30 simple pores in each ventral submarginal line between lateral stigmatic areas................................................................................................ C. marika sp. n.
Fiji is about 1500 miles (2400 km) north of New Zealand. Within New Zealand, Crystallotesta species have been found on 13 plant families but most commonly on Myrtaceae and Nothofagaceae ; there are no records off Polypodiaceae . The discovery of a species in a genus otherwise thought to be restricted to New Zealand was surprising and begs the question as to how it might have arrived in Fiji. Obviously it is possible that C. marika does occur in New Zealand (but has yet to be discovered there) and has been accidentally introduced into Viti Levu at some time in the fairly recent past. However, it is at least as likely that it arrived a long time ago, before the arrival of man (about 4000 years ago). New Zealand is believed to have been part of the supercontinent Gondwana but became separated from Australia and New Caledonia more than 80 million years ago. Since then it has had a chequered history but has never been connected with the Oceania ecozone, to which Fiji belongs. Thus, despite the unmistakable Gondwanan influences in the flora of Viti Levu today, there is no geological evidence to support the idea that it was once part of Gondwana. However, it is considered ( Yan & Kroenke, 1993) that part of the Norfolk Ridge – a part of Gondwana that also included New Caledonia – became separated and moved into the South Fiji Basin. Yan and Kroenke (1993) further suggest that a fragment of this ridge, represented by the Tongan island of ‘Eua, moved eastward and eventually became incorporated into the Tongan island group. In order to account for the strong Gondwanan influences in the fauna and flora of Fiji, it has been suggested ( Mueller-Dombois & Fosberg, 1998) that ‘Eua came close enough to Viti Levu to allow transfer of some of its distinct flora. If this scenario is correct, then perhaps the ancestor of C. marika was carried on ‘Eua and also managed to transfer to the nascent Viti Levu, along with some of the flora. If so, then it is quite possible that further species in this genus will be discovered on other south Pacific island groups, particularly ‘Eua!
Name derivation. The specific name marika is in honour of Dr. Marika Tuiwawa, Curator of the South Pacific Regional Herbarium, Institute of Applied Sciences, University of South Pacific, Suva, who was extremely helpful during the visit by CJH when these specimens were collected.
| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |