
Callinera emiliae, Kajihara, Hiroshi, 2007
|
publication ID |
https://doi.org/10.5281/zenodo.176314 |
|
DOI |
https://doi.org/10.5281/zenodo.6239995 |
|
persistent identifier |
https://treatment.plazi.org/id/03A487F3-FFEF-FFCD-FF6D-3B197C80FA2C |
|
treatment provided by |
Plazi |
|
scientific name |
Callinera emiliae |
| status |
sp. nov. |
Callinera emiliae sp. nov. ( Figs 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 )
Diagnosis: A Callinera having lateral sensory organs; sub-epidermal glandular cells absent; blood vascular system without cephalic ventral connective; nervous system with double dorsal and single ventral cerebral commissures; foregut nerves fusing to form ganglion in front of mouth; epidermis with constrictions in intestinal region; and intestinal sphincters present.
Etymology: The specific name is dedicated to Dr Emilia S. Yap, a Filipino food scientist who guided my collecting trip in the Philippines.
Material examined: Holotype, ZIHU-3187, male, serial transverse sections of complete specimen, 77 slides. Paratypes: UPLB-MNH-Z-NS-0389, serial transverse sections of the anterior region of the body, 27 slides; UPLB-MNH-Z-NS-0390, serial transverse sections of the anterior region of the body, 7 slides.
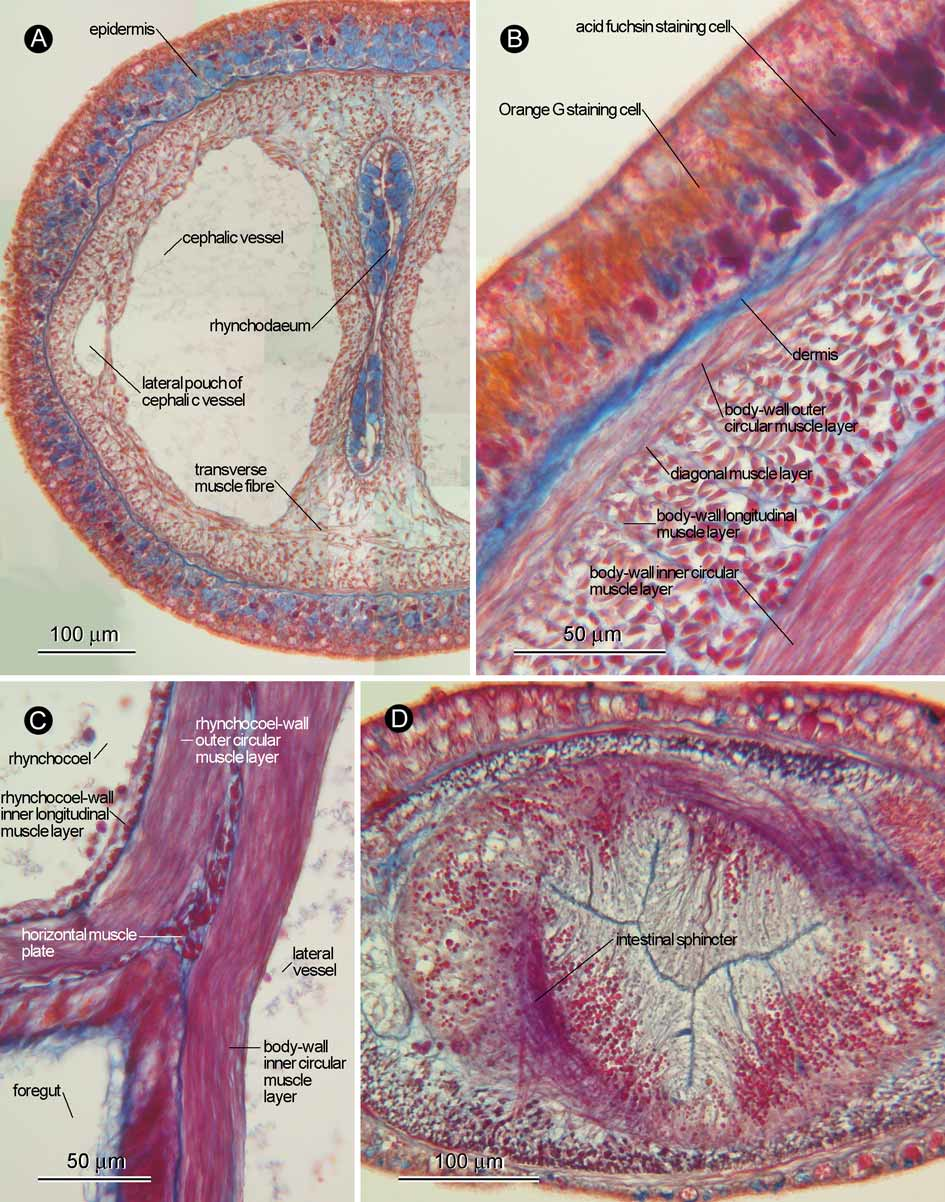
External features: Body about 7 cm long and 1.8 mm wide when anaesthetized ( Fig. 2 View FIGURE 2 A). Head translucent, dorsoventrally flattened, and demarcated from succeeding body; white rhynchodaeum visible through cephalic surface; with neither eyes nor cephalic furrows ( Fig. 2 View FIGURE 2 B). Foregut region cross-sectionally rounded; foregut cream-white. Epidermis translucent except in posterior foregut region, where epidermal hue suddenly changing into light flesh colour; from there on backward, epidermis gradually resuming translucent appearance; pair of lateral organs present, recognized as horizontally elongated epidermal indentation, located on lateral side of body on each side just posterior to light flesh colour epidermal band ( Fig. 2 View FIGURE 2 C). Proboscis white in colour. In intestinal region six constrictions present ( Fig. 2 View FIGURE 2 A, D); gonads arranged in row on each side.
Body wall, musculature, and parenchyma: Ciliated epidermis up to 100 µm thick in brain region, gradually thinner posteriorly, 5–30 µm thick in intestinal region; basophilic glandular cells predominant in pre-cerebral region ( Fig. 3 View FIGURE 3 A); in foregut region, acid fuchsin staining glandular cells predominant before being abruptly replaced by Orange G staining acidophilic cells in flesh-coloured epidermal band ( Fig. 3 View FIGURE 3 B); posteriorly, epidermis containing basophilic and neutrophilic glandular cells, in addition to acidophilic glandular cells; E ( b) = 0.08 ( holotype ZIHU-3187 and paratype UPLB-MNH-Z-NS-0389) and 0.06 ( paratype UPLB- MNH-Z-NS-0390); E ( i) = 0.04 ( holotype). Dermis up to 10 µm thick in foregut region; processes of connective tissue into epidermis not found; innermost side of dermis not forming mesh-like structure. Body-wall musculature composed of outer circular and inner longitudinal muscle layers; inner circular muscle layer present ( Fig. 3 View FIGURE 3 B) from mouth through rhynchocoel posterior chamber; thin diagonal layer present between outer circular and longitudinal muscle layers ( Fig. 3 View FIGURE 3 B). Pre-cerebrally, transverse muscle fibres running below rhynchodaeum and cephalic vessels ( Fig. 3 View FIGURE 3 A). Longitudinal muscle plate present between rhynchocoel and alimentary canal ( Fig. 3 View FIGURE 3 C), terminating before rhynchocoel posterior chamber. Intestine with six sphincters ( Fig. 3 View FIGURE 3 D), corresponding with epidermal constrictions.
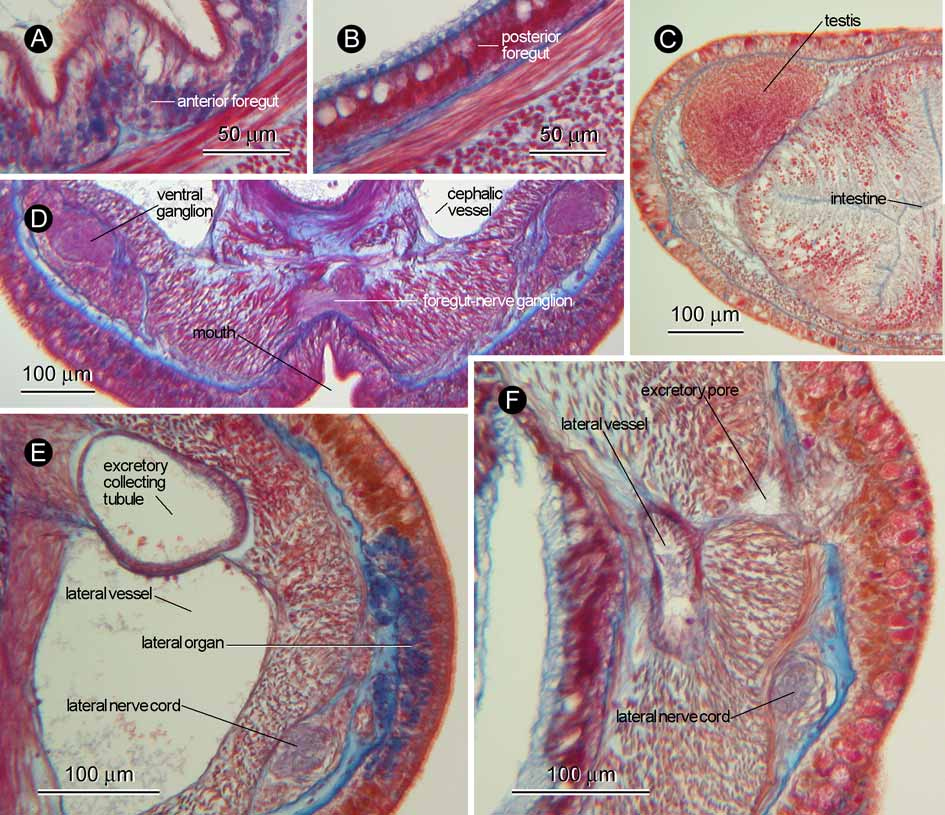
Proboscis apparatus: Proboscis pore opening mid-ventrally near tip of head, leading to rhynchodaeum; rhynchodaeal wall containing basophilic glandular cells ( Fig. 3 View FIGURE 3 A) except just in front of proboscis insertion ( Fig. 4 View FIGURE 4 A). Rhynchodaeal sphincter present just in front of proboscis insertion, composed of circular muscles derived from body-wall longitudinal muscle layer ( Fig. 4 View FIGURE 4 A, B). Proboscis insertion situated near brain ring, composed of fibres from rhynchodaeal sphincter; pair of proboscis nerves originating from ventral ganglia, running below cephalic vessel and entering proboscis via insertion ( Fig. 4 View FIGURE 4 C). Rhynchocoel extending about 60% of body length, with wall composed of outer circular and inner longitudinal muscle layers ( Fig. 3 View FIGURE 3 C). Rhynchocoel posterior chamber (or ‘muscular sac’) present, its wall mid-dorsally derived from body-wall inner and outer circular muscle layers; pair of ‘pleats’ present between anterior and posterior rhynchocoel junction, also originating from body-wall inner and outer circular muscle layers ( Fig. 4 View FIGURE 4 D–K). Proboscis histologically differentiated into four regions: first region [= Bergendal’s region ( Kajihara 2006)], 160 µm long, composed of glandular epithelium and longitudinal muscle layer, pair of proboscis nerves present between these two layers ( Fig. 5 View FIGURE 5 A); second region, 720 µm long, possessing circular muscle layer between glandular epithelium and longitudinal muscle layer, with pair of proboscis nerves situated between glandular epithelium and circular muscle layer ( Fig. 5 View FIGURE 5 B), glandular epithelium posteriorly becoming thicker and densely ciliated ( Fig. 5 View FIGURE 5 C); third region, 1.2 mm long, with glandular epithelium containing distinct acidophilic glandular cell mass above proboscis nerves ( Fig. 5 View FIGURE 5 D); fourth main region, probably accounting for more than 80% of entire proboscis length, possessing epithelium basically composed of cells with neutrophilic cytoplasm, containing acidophilic and basophilic granules ( Fig. 5 View FIGURE 5 E), followed by proboscis retractor muscle ( Fig. 5 View FIGURE 5 F); fourth main region and proboscis retractor muscle extending into rhynchocoel posterior chamber ( Fig. 5 View FIGURE 5 F).
Alimentary canal: Mouth opening just posterior to brain; foregut histologically differentiated into two regions: anterior region possessing epithelium composed of ciliated glandular cells containing basally basophilic and distally neutrophilic contents ( Fig. 6 View FIGURE 6 A); posterior region possessing epithelium dominated by acidophilic glandular cells, sporadically containing neutrophilic cells ( Fig. 6 View FIGURE 6 B). No intestinal caecum. Main intestinal canal possessing epithelium composed of tall columnar cells containing neutrophilic cytoplasm with basal acidophilic granules ( Fig. 6 View FIGURE 6 C); intestine without lateral diverticula.
Blood system: Pair of cephalic vessels meeting anteriorly, dorsoventrally pierced by muscle strands; in rhynchodaeal region, dorsoventral muscle strands more or less sagittally continuous, making cephalic vessel seemingly possess lateral pouch ( Fig. 3 View FIGURE 3 A); with neither dorsal nor ventral connectives. Post-cerebrally, lateral vessels first situated inside body-wall inner circular muscle layer; posteriorly, near junction between anterior and posterior foregut regions, lateral vessels traversing through body-wall inner circular muscle layer to be situated immediately outside it. Mid-dorsal vessel absent.
Nervous system: Brain and lateral nerves situated between dermis and body-wall outer circular muscle layer; inner neurilemma incompletely developed, outer neurilemma well developed ( Fig. 4 View FIGURE 4 C). Two dorsal and one ventral cerebral commissures present. Pair of foregut nerves anteriorly meeting to form ganglion just before mouth ( Fig. 6 View FIGURE 6 D). Mid-dorsal nerve present between dermis and body-wall outer circular muscle layer, frequently sending fibres downward to rhynchocoel wall.
Sense organs: Pair of lateral organs situated anterior to rhynchocoel posterior chamber, containing basophilic cells ( Fig. 6 View FIGURE 6 E). Neither cerebral sense organs nor statoliths present. No eyes.
Excretory system: Excretory system composed of thin walled collecting tubule on each side of body above lateral blood vessel ( Fig. 6 View FIGURE 6 E), opening to exterior at its posterior end ( Fig. 6 View FIGURE 6 F). In holotype, anterior glandular mass not well developed; anterior end of excretory system not appearing to enter lateral vessel. In one of two paratypes (UPLB-MNH-Z-NS-0389), anterior end of excretory collecting tubule entering lateral blood vessel; the other paratype (UPLB-MNH-Z-NS-0390) lacking posterior portion of body containing excretory system.
Reproductive system: The holotype was a male. Sexes of paratypes are unknown. Testes up to 200 µm in diameter, arranged in row above lateral blood vessel on each side ( Fig. 6 View FIGURE 6 C).
Remarks: Callinera emiliae sp. nov. is identified as the member of the genus Callinera by possessing a nervous system situated between the dermis and body-wall outer circular muscle layer, the rhynchocoel posterior chamber (= muscular sac), and no cerebral sense organs. Callinera emiliae sp. nov. can be distinguished from the congeners by the characteristics summarized in Table 1.
The morphology of the rhynchocoel posterior chamber in Callinera emiliae sp. nov. was most clearly distinguishable in one of the two paratypes that had lost the proboscis. There are thin pleats originating from the body-wall inner and outer circular muscle layers in the junction between the anterior and posterior rhynchocoel chambers. In the holotype, however, the pleats were pressed against the rhynchocoel wall by the proboscis, which made tracing of the morphology difficult. Remarkably, the structure of the rhynchocoel posterior chamber is similar to that reported in Tubulanus borealis Friedrich, 1936 . The rhynchocoel posterior chamber is regarded as the possible synapomorphy for the genus Callinera in the cladistic analysis based on 50 morphological characters performed for 49 palaeonemerteans by Sundberg and Hylbom (1994). However, T. borealis was not included in their analysis due to the plethora of unknown character states in this taxon. Anyhow, the two genera Callinera and Tubulanus appear to be not clearly discriminated by the traditionally used characters, viz., the presence/absence of the cerebral sensory organs and rhynchocoel posterior chamber.
TABLE 1. Comparison of six characters among Callinera species. Data compiled from Bergendal (1900a, b, c, 1901, 1903), Rogers et al. (1992), Gibson and Sundberg (1999), Senz (2000), Chernyshev (2002), and Kajihara (2006).
Characters and character states:
A: Lateral sensory organs: (0) absent; (1) present.
B: Body-wall longitudinal muscle layer: (0) without; (1) with sub-epidermal glandular cells. C: Proboscis: (0) without; (1) with circular muscle layer in region following Bergendal’s region. D: Cephalic blood vascular system: (0) without; (1) with ventral lacuna. E: Dorsal cerebral commissure(s): (1) one; (2) two.
F: Foregut nerves: (0) fuse to form a single median nerve before branching anterior to mouth, or remaining as two distinct nerves (1).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |