
Obelia bidentata S.F. Clark, 1875
|
publication ID |
https://doi.org/ 10.5281/zenodo.556851 |
|
publication LSID |
lsid:zoobank.org:pub:985C0239-D00C-457D-B593-76A3081BCEEA |
|
DOI |
https://doi.org/10.5281/zenodo.6015998 |
|
persistent identifier |
https://treatment.plazi.org/id/03A787C7-4904-FFAB-FF58-FECCFBBCFC39 |
|
treatment provided by |
Plazi |
|
scientific name |
Obelia bidentata S.F. Clark, 1875 |
| status |
|
Obelia bidentata S.F. Clark, 1875 View in CoL
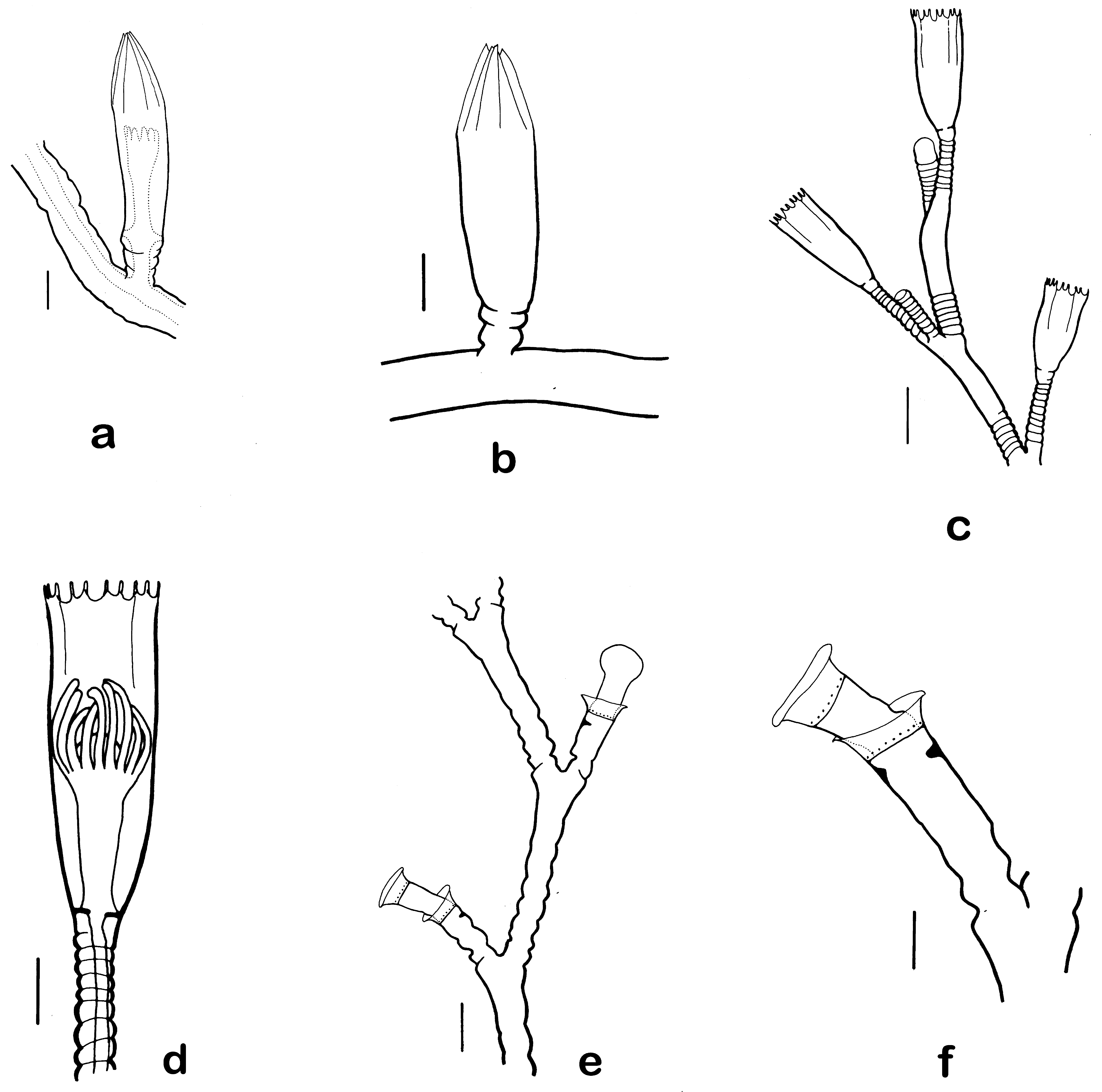
Figs. 17 View FIGURE 17 c, d
Obelia bidentata S.F. Clark, 1875: 58 View in CoL , pl. 9, fig. 2.
Type locality. USA: New York, Long Island, Greenport , on wharf piles at low water ( S.F. Clark 1875: 59).
Material examined. NS: Minas Basin, Kingsport, low intertidal, on bryozoan and hydroid stems on a stone, 20.viii.1970, one young colony, up to 1 cm high, without gonophores, coll. K.W. Petersen, SNM HYD-001061.— NS: Minas Basin, Medford Beach near Kingsport, on sandy silt near low water, 28.viii.1984, five colonies, up to 7.5 cm high, without gonophores, coll. J.S. Bleakney, ROMIZ B507.—NS: Minas Basin, Kingsport, low tide, --. ix.1999, underside of sandstone overhang, two colonies or colony fragments, up to 5.4 cm high, without gonophores, coll. J.S. Bleakney, ROMIZ B3121.
Description. Colonies erect, bottle-brush-shaped, up to 7.5 cm high, arising from a creeping hydrorhiza (SNM HYD-001061; ROMIZ B3121) or from a root-like mass of tangled stolons with adhering fine sediments (ROMIZ B507). Hydrocaulus single and not forked, straight to slightly geniculate, bearing regular branches, strongly polysiphonic basally, gradually becoming thinner, with monosiphonic distal end. Central axis of hydrocaulus divided into long internodes, each internode annulated basally, smooth elsewhere, distal end with large apophysis supporting bases of two branches as well as a single pedicel terminating in a hydranth. Perisarc of moderate thickness. Branches of hydrocaulus at proximal end of colony sometimes fascicled basally but usually monosiphonic, given off on alternate sides of central axis of hydrocaulus, usually with a pair of branches originating very close together, these diverging from each other at an angle of about 90 degrees; two closest branch pairs arising on opposite sides of hydrocaulus, the two pairs thereby sending out branches into four quadrants around hydrocaulus; branch internodes annulated basally, smooth elsewhere. Primary branches quite short, up to 0.8 cm long, each either unbranched and giving off alternate pedicels or more often alternately branched; secondary branches very short, directed upwards and outwards, bearing 2-4 alternate pedicels with terminal hydrothecae; in addition, a single pedicel with terminal hydrotheca usually given off near base of each secondary branch and directed laterally. Pedicels of varied length within a single colony, either annulated throughout or smooth in middle and annulated proximally and distally, supporting a hydrotheca at terminal end. Hydrothecae deeply campanulate, about 0.45–0.55 mm long, 0.18–0.22 mm at margin, 0.7–0.8 mm wide at diaphragm, tapering basally, slightly flaring at distal end, adcauline wall at basal end somewhat more convex than abcauline wall. Vertical pleats extending part-way down hydrothecal wall from incision between adjoining marginal cusps. Hydrothecal margin with about 10–12 cusps separated by U-shaped incisions, each cusp with two sharp-pointed lateral keels extending into hydrothecal cavity. Hydrothecal diaphragm thin, appearing straight to slightly oblique, basal chamber fairly large, cup-shaped.
Gonophores and gonothecae not seen.
Cnidome
England ( Östman 1999)—pseudo-microbasic b-mastigophores, sB b -type: 8.0–9.0 µm x 2.5–3.0 µm (undischarged)
Brazil ( Migotto 1996; Östman 1999)—
pseudo-microbasic b-mastigophores, A-type: 6.0–6.5 µm x 2.0–2.5 µm (undischarged)
isorhizas (I D -type?): 5.0–5.5 µm x 2 µm (undischarged)
isorhizas (I d - type?): 5.0–6.5 µm x 1.0–1.5 µm (undischarged)
Remarks. Obelia bidentata S.F. Clark, 1875 and O. bicuspidata S.F. Clark, 1875 have long been regarded as conspecific (e.g., Jäderholm 1903; Fraser 1912, 1944; Nutting 1915; Cornelius 1975, 1982, 1990, 1995b; Calder 1991; Medel & Vervoort 2000). Of the two simultaneously established names, precedence was assigned to O. bidentata by Jäderholm (1903), acting as First Reviser ( ICZN Art. 24.2). His selection was widely overlooked, with usage of the name O. bicuspidata prevailing for much of the 20th century, until correct nomenclature of the species was restored by Cornelius (1975). Since then, the valid name O. bidentata has gained near universal adoption. Both nominal species were originally described from Long Island Sound, on the Atlantic coast of the United States, by S.F. Clark (1875). The type locality of O. bidentata is Greenport, New York, while that of O. bicuspidata is nearby at the Thimble Islands , Connecticut. S.F. Clark differentiated the two based on differences in colony habit, in the relative diameter and depth of their hydrothecae, and in the length of their hydrothecal pedicels, but such differences have been widely viewed as taxonomically insignificant. An extensive synonymy list of the species was given by Medel & Vervoort (2000).
While there is little reason to suspect that Obelia bidentata and O. bicuspidata are distinct, the possibility exists that multiple species may have been combined under the currently accepted name since S.F. Clark’s (1875) original account. Obelia bidentata , now reported to be essentially cosmopolitan ( Medel & Vervoort 2000; Peña Cantero & Gili 2006), seems more likely to be a largely temperate to warm-temperate species with a more restricted distribution than presently assumed. A population of hydroids meeting the currently accepted broad concept of this species from warm waters off the east cost of Florida, USA, has already been treated as distinct (Calder 2013: 59). Although hydrothecae of specimens from locations including the Caribbean Sea ( Vervoort 1968), Bermuda ( Calder 1991), and Brazil ( Migotto 1996) appear essentially identical in morphology with those examined here, certain hydroids from those tropical and subtropical areas tend to be small, unbranched or irregularly branched, and with monosiphonic or weakly polysiphonic hydrocauli rather than being large (5–10 cm or more), extensively and regularly branched, and with strongly polysiphonic hydrocauli as in populations from temperate waters. Such differences are usually dismissed as merely phenotypic variations within the species. Of possible taxonomic significance, however, is the discovery by Östman (1999) of cnidome differences in hydroids identified as O. bidentata from Brazil (with small monosiphonic colonies) and England (with large polysiphonic colonies). As for possible molecular evidence of genetic distance in populations assigned to the species, see remarks given two paragraphs below. Meanwhile, on the basis of biogeography and apparent temperature tolerances of the species, records of O. bidentata from the Antarctic Ocean (reviewed in Peña Cantero & Gili 2006) seem open to reconsideration. In the northern hemisphere, this hydroid is primarily a temperate zone species that does not penetrate far into boreal waters ( Fraser 1944; Cornelius 1988, 1990, 1995b; this study), and it is completely absent in subarctic and Arctic regions ( Naumov 1960; Calder 1970; Schuchert 2001a; Antsulevich 1915). Also worthy of further investigation is the identity of a population of hydroids resembling O. bidentata , but having very small colonies (<1 cm) and small hydrothecae, that inhabits oligohaline areas at the head of estuaries on the Atlantic coast of the United States. Similar colonies have been observed in the upper reaches of a small tropical estuary in Brazil ( Calder & Maÿal 1998). The species was reported across a salinity range from 0.5–34‰ in estuaries of South Carolina ( Calder 1976), but the identity of hydroids occurring in areas of very low salinity needs to be reexamined.
Specimens examined here from Minas Basin (SNM HYD-001061, ROMIZ B507, ROMIZ B3121) correspond closely with the original account of Obelia bidentata by S.F. Clark (1875). In addition to the identical morphology of their hydrothecae, two of the colonies were large (reaching more than 7 cm high) and had strongly polysiphonic stems. In having branches extending outwards from four sides of the stem, the colonies are somewhat similar in overall appearance to those of Hartlaubella gelatinosa ( Pallas, 1766) , a species also recorded from the Bay of Fundy. That resemblance has already been noted by Cornelius (1982, 1995b). Trophosomes of those two species can be readily distinguished by the shape of the lateral spines on cusps of their hydrothecae. Those of O. bidentata are slender, elongate, keel-shaped, and separated by U-shaped incisions, whereas those of H. gelatinosa are blunt, triangular, and separated by V-shaped incisions. In addition, gonophores of O. bidentata are free medusae while those of H. gelatinosa are fixed sporosacs. Gonophores and gonothecae were absent in colonies examined during this study, but distinctive trophosomal characters leave no doubt about their identity.
Molecular studies based on specimens from northwestern Europe and North Carolina (e.g., Govindarajan et al. 2006; Leclère et al. 2009; Peña Cantero et al. 2010; Maronna et al. 2016) have yielded conflicting and confusing results about the affinities of O. bidentata . Analyses have suggested that the species occupies a clade that is quite distant from others assigned to the genus Obelia Péron & Lesueur, 1810 . Its inclusion in Obelia is nevertheless supported by the morphology of the medusa stage, which is typical of the genus ( Nutting 1915; Hamond 1957; Cornelius 1982). Most phylograms to date have included the species as part of a cluster also comprising Laomedea flexuosa Alder 1857 , Gonothyraea inornata Nutting, 1901b , and Obelia longissima ( Pallas, 1766) . Meanwhile, hydroids attributed to O. bidentata from Europe and North America appear genetically quite distant in the analyses, possibly lending further support to the idea that a species complex exists under the name. More molecular work is needed on hydroids ascribed to O. bidentata , ideally including specimens collected near its type locality and conforming with the original description of S.F. Clark (1875).
In Long Island Sound, S.F. Clark (1875) found O. bidentata “…in considerable abundance…” on 0 5 August 1874, while specimens referred by him to O. bicuspidata were collected 23 September 1874. Specimens in this study from Minas Basin ( ROMIZ B507; ROMIZ B3121) were collected during the same months, at a time of year when water temperatures are still relatively warm in the area. In estuaries of South Carolina, the species is active for much of the year except for a few weeks during the coldest part of the winter, when water temperatures drop to 10° C or below ( Calder, 1990). Coinciding with such hibernal conditions, colonies become dormant. As with colonies examined here, S.F. Clark found his specimens at shallow depths [ O. bidentata on wharf pilings at low water; O. bicuspidata on “reefs” at 3–5 fm (5–9 m)]. Similar hydroids were described from the intertidal zone in pools at Holme-next-the-Sea near Hunstanton , Norfolk, UK, by Cornelius (1988). Illustrations of a complete young colony of the species in Cornelius (1990: 548, fig. 1a; 1995b: 293, fig. 68A) were based on a specimen from that area. Hydroids from Minas Basin are similar in colony form. One lot of those specimens ( ROMIZ B507), taken at low water on a flat of fine sediments, were anchored to the soft substrate by a root-like mass of tangled stolons. This type of hydrorhiza was also observed on colonies from sandy substrates by Cornelius (1975: 261).
As with Hydractinia symbiolongicarpus Buss & Yund, 1989 , discussed above, the occurrence of O. bidentata in Minas Basin appears to be disjunct in geographic distribution. It is not known from elsewhere in the Bay of Fundy or from any other location in Atlantic Canada. The centre of distribution of the species along the east coast of North America is in warmer waters south of Cape Cod . The nearest reported population of O. bidentata , from Casco Bay , Maine ( Fraser 1944, as O. bicuspidata ), appears to be disjunct as well.
Recorded distribution. Bay of Fundy: recorded for the first time.
Eastern North America: Minas Basin, NS (reported herein) to the Caribbean Sea ( Galea 2010b).
Elsewhere: Worldwide, tropical and temperate waters ( Medel & Vervoort 2000).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Obelia bidentata S.F. Clark, 1875
| Calder, Dale R. 2017 |
Obelia bidentata
| Clark 1875: 58 |