
Nectoneanthes uchiwa, Sato, 2013
|
publication ID |
https://doi.org/ 10.1080/00222933.2012.743609 |
|
DOI |
https://doi.org/10.5281/zenodo.4742637 |
|
persistent identifier |
https://treatment.plazi.org/id/03A8316D-FFD4-C829-F1F6-CC6FF45DF988 |
|
treatment provided by |
Carolina |
|
scientific name |
Nectoneanthes uchiwa |
| status |
sp. nov. |
Nectoneanthes uchiwa View in CoL sp. nov.
(Japanese name: Uchiwa-gokai) (Korean name: Neob-jeok-bal-charm-gaet-ji-reong-i)
( Figures 4A View Figure 4 , 9–14 View Figure 9 View Figure 10 View Figure 11 View Figure 12 View Figure 13 View Figure 14 )
Nereis oxypoda: Izuka, 1912: 171–173 View in CoL , pl. 18, figs. 8–11 (in part); Mori et al., 1932: 3, pl. 1, fig. 2, pl. 3, figs. 1–6 (in part); Okuda, 1933: 247, pl. 13, figs. f–h; Monro, 1934: 362–363 (in part, 2 specimens collected by Ping).
Nereis (Neanthes) oxypoda: Khlebovich, 1963: 55 View in CoL .
Neanthes oxypoda: Imajima and Hartman, 1964: 145 View in CoL .
Neanthes (Nectoneanthes) oxypoda View in CoL var.: Wilson, 1984: 220.
Neanthes View in CoL sp.: Wilson, 1988: 7–8.
Nectoneanthes oxypoda: Imajima, 1972: 113–118 View in CoL , figs. 35 b, c (in part, atokes); Paik, 1972: 135, fig. 4; Paik, 1973: 82–84, figs. 1k–l, 2; Wu et al., 1985: 164–167, figs. 92–93; Yang and Sun, 1988: 39, figs. 8A–D; Imajima, 1996: 144–145, figs. 116 b, c (in part, atokes); Khlebovich, 1996: 115–116, pl. XXII; Lee et al., 2003: 189, 190; Yamanishi and Sato, 2007: 183.
Nectoneanthes oxypoda sensu Imajima, 1972 View in CoL : Hanafiah et al., 2006: 209; Saito et al., 2011: 47.
Type material
Holotype ( NSMT Pol H-549, Figure 10A, B View Figure 10 ), atokous individual, collected from off Hachihama (34 ◦ 07 ′ 20 ′′ N, 133 ◦ 07 ′ 20 ′′ E) in Kojima Bay in Seto Inland Sea , Okayama Prefecture on 17 December 1906, by A. Izuka (the original estuarine habitat of this site was changed to dry land and a freshwater pond before 1959 by reclamation projects) . Paratype ( NSMT Pol P-550), 13 atokous individuals, data as for holotype . Holotype and paratypes deposited in polychaete collection in Tsukuba Research Departments of National Museum of Nature and Science, Tsukuba, Ibaraki Prefecture, Japan .
Other material examined
Atokes collected from Japan. Seto Inland Sea: Hiroe-gawa River (33 ◦ 07 ′ 32 ′′ N, 133 ◦ 07 ′ 4 ′′ E), Imazaike , Mitsuya , Nyugawa , Ehime Prefecture, 31 August 1969, 2 (BW, 1.3–1.7 mm; NSMT Pol 18532–18533) . Intertidal flats at Nagai-hama (33 ◦ 07 ′ 10 ′′ N, 131 ◦ 07 ′ 15 ′′ E), Yukuhashi , Fukuoka Prefecture, 26 October 2003, coll. Y. Henmi et al., 1 (BW, 2.3 mm; OMNH) . Intertidal flats at Sone-higata (33 ◦ 07 ′ 14.4 ′′ N, 130 ◦ 07 ′ 29.5 ′′ E), Kitakyushu , Fukuoka Prefecture, 25 April 2002, coll. Y. Henmi et al., 1 (BW, 3.9 mm; OMNH) . Ariake Sea: Sta. 50 (32 ◦ 33.2 ′ N, 130 ◦ 19.3 ′ E, 20 m deep), September 1958, coll. Seikai National Fisheries Research Institute, 5 (BW, 1.0 mm; NSMT Pol 5280–5284) . Intertidal flats at Kojiro-nagahama (32 ◦ 07 ′ 19 ′′ N, 130 ◦ 07 ′ 42 ′′ E), Kunimi , Nagasaki Prefecture, Isahaya Bay , 19 October 1994, coll. H. Yamashita, 1 (BW, 3.5 mm; ZMH P-26036) . Intertidal flats at Isaki (32 ◦ 07 ′ 45 ′′ N, 130 ◦ 07 ′ 00 ′′ E), Konagai , Nagasaki Prefecture, Isahaya Bay , 17 March 1995, coll. M. Sato, 1 (BW, 2.3 mm; ZMUC-POL-2220) . Intertidal flats at the mouth of Tagorigawa River (32 ◦ 07 ′ 13 ′′ N, 130 ◦ 07 ′ 46–47 ′′ E), Takesaki , Saga Prefecture, 7 March 2004, coll. Y. Henmi et al., 2 (BW, 3.0– 4.5 mm; OMNH) . Intertidal flats at the mouth of Shiota-gawa River (33 ◦ 07 ′ 50 ′′ N, 130 ◦ 07 ′ 10 ′′ E), Kashima , Saga Prefecture, 19 April 2004, coll. Y. Henmi et al., 1 (BW, 2.0 mm; OMNH) . Intertidal flats off Yanagawa (33 ◦ 07 ′ 21 ′′ N, 130 ◦ 07 ′ 03 ′′ E), Fukuoka Prefecture, 10 May 1998, coll. M. Sato et al., 2 (BW, 2.5–3.0 mm; MS); 27 April 2002, coll. Y. Henmi et al., 1 (BW, 2.2 mm; OMNH) . Intertidal flats at the mouth of Yabe-gawa River (33 ◦ 07 ′ 30 ′′ N, 130 ◦ 07 ′ 07 ′′ E), Fukuoka Prefecture, 30 April 2002, coll. Y. Henmi et al., 1 (BW, 2.0 mm; OMNH) . Intertidal flats in Omuta-gawa River (33 ◦ 07 ′ 13 ′′ N, 130 ◦ 07 ′ 02 ′′ E), Fukuoka Prefecture, 17 April 2003, coll. M. Sato, 1 (BW, 1.5 mm; MS); 16 April 2004, coll. M. Sato et al., 5 (BW, 1.5–4.0 mm; NSMT Pol 11413) . Intertidal flats at Arao (32 ◦ 07 ′ 17 ′′ N, 130 ◦ 07 ′ 32 ′′ E), Kumamoto Prefecture, 2 September 2008, coll. H. Tsutsumi, 5 (BW, 2.1–3.0 mm; NSMT Pol 11414) . Intertidal flats at the mouth of Kikuchi-gawa River (32 ◦ 07 ′ 28 ′′ N, 130 ◦ 07 ′ 02 ′′ E), Tamana , Kumamoto Prefecture, 22 April 2004, coll. Y. Henmi et al., 1 (BW, 3.0 mm; OMNH) . Intertidal flats at the mouth of Midori-kawa River (32 ◦ 07 ′ 55 ′′ N, 130 ◦ 07 ′ 40 ′′ E), Kumamoto Prefecture, 1 September 2008, coll. H. Tsutsumi, 1 (BW, 3.0 mm; NSMT Pol 11408) . Intertidal flats at Aitsu (32 ◦ 07 ′ 22 ′′ N, 130 ◦ 07 ′ 28 ′′ E), Amakusa ( Marine Biological Station Kumamoto University ), Kumamoto Prefecture, 15 March 2002, coll. M. Sato et al., 3 (BW, 2.5–3.2 mm; NSMT Pol 11412) . Shiranui Sea (Yatsushiro Sea): Intertidal flats at the mouth of Hikawa River (32 ◦ 07 ′ 17 ′′ N, 130 ◦ 07 ′ 59 ′′ E), Kumamoto Prefecture, 16 September 2004, coll. Y. Henmi et al., 2 (BW, 2.6–3.0 mm; OMNH) .
Atokes collected from Korea. Hampyong Bay : Sta. 73 (35 ◦ 07 ′ 06 ′′ N, 126 ◦ 07 ′ 16 ′′ E, 5 m deep) and Sta. 86 (35 ◦ 07 ′ 40 ′′ N, 126 ◦ 07 ′ 50 ′′ E, 5 m deep), June 2008, coll. J.-W. Choi, 4 (BW, 1.0–3.0 mm; KIOST) . Beolkyo Bay : Sta. 18 (34 ◦ 07 ′ 37 ′′ N, 127 ◦ 07 ′ 45 ′′ E, 5 m deep), February 2009, coll. J.-W. Choi, 15 (BW, 1.1–4.3 mm; NSMT Pol 11411; KIOST) . Gwangyang Bay : Sta. 5 (34 ◦ 07 ′ 55 ′′ N, 127 ◦ 07 ′ 29 ′′ E, 10 m deep), 7 August 2008, coll. J.-W. Choi, 1 (BW, 2.2 mm; KIOST) ; November 2010, coll. J.-W. Choi, 1 (BW, 3.0 mm; KIOST) . Sta. 6 (34 ◦ 07 ′ 58 ′′ N, 127 ◦ 07 ′ 53 ′′ E, 10 m deep), 7 August 2008, coll. J.-W. Choi, 4 (BW, 1.5–2.3 mm; KIOST) .
Epitokes collected during reproductive swarming in Japan. Omuta-gawa River (33 ◦ 07 ′ 15 ′′ N, 130 ◦ 07 ′ 08 ′′ E), Fukuoka Prefecture, 20 April 2004 (22:30–23:00), coll. H. Nakashima, 1 female ( NSMT Pol 11409); 8 May 2004 (0:54–1:25), coll. H. Nakashima, 3 males ( NSMT Pol 11410, MS) .
Diagnosis
Broad continuous band of many paragnaths (more than 200) present in dorsal and ventral areas of oral ring of proboscis, covering groups V and VII–VIII, except for group VI remaining in compact clusters separated from other paragnaths. Relatively large number of paragnaths in group II (14–43 on each side), III (5–34) and IV (17–49 on each side) in maxillary ring of proboscis.
Description
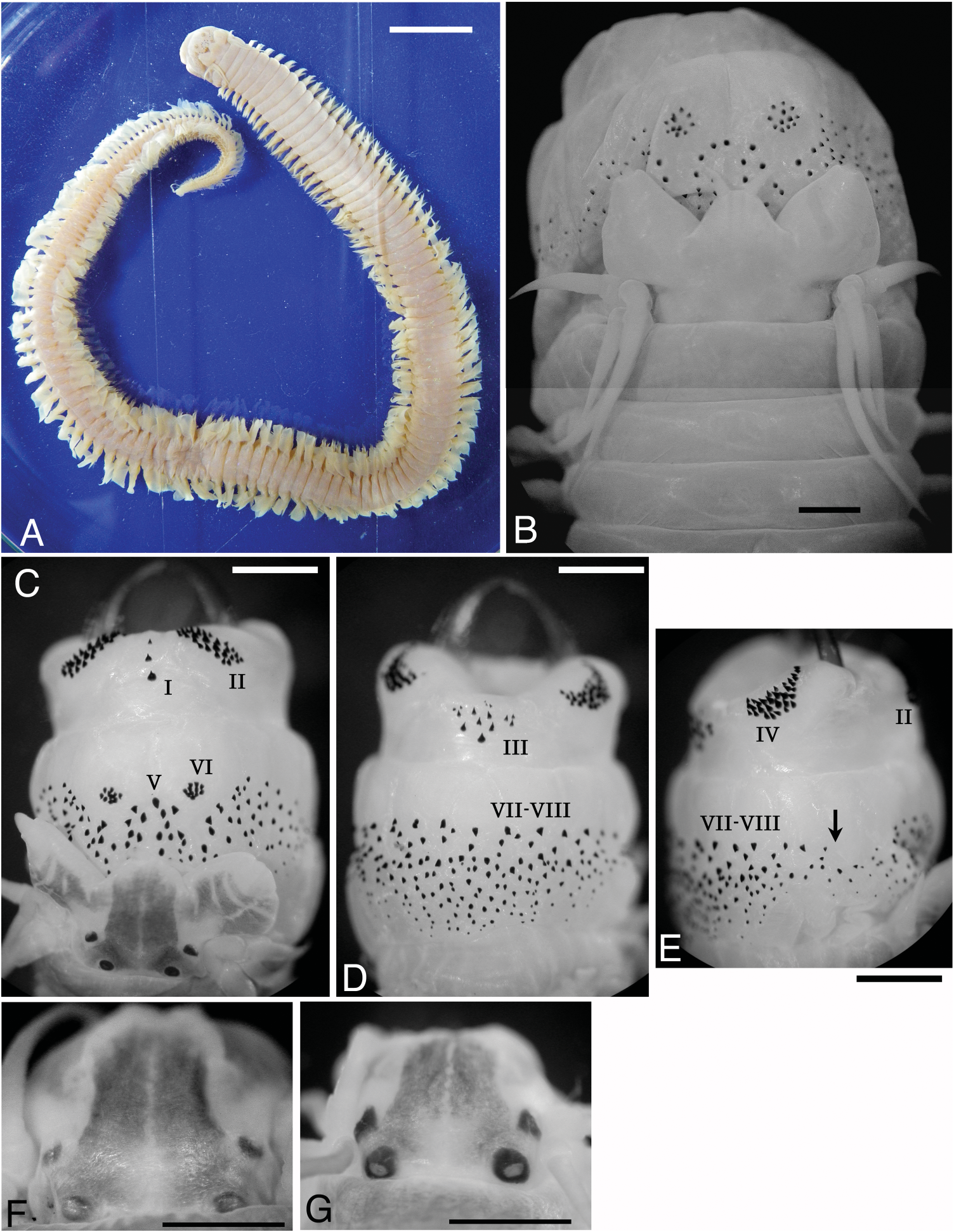
Type and non-type atokes. Holotype, complete individual 170 mm BL, 5.8 mm BW, with 154 chaetigers, with proboscis fully everted ( Figure 10A, B View Figure 10 ). Paratypes 145–260 mm BL, 3.7–6.0 mm BW; largest complete specimen 260 mm BL, 6.0 mm BW, with 185 chaetigers. Non-types (n = 61), 1.0– 5.5 mm BW.
Body stout anteriorly, posteriorly gradually tapering toward pygidium ( Figure 10A View Figure 10 ). Dorsum convex, venter relatively flat with longitudinal midventral groove. Colour in preserved specimens whitish cream with brownish pigmentation in anterior dorsum.
Prostomium with trapezoidal shape, broader than long, with pair of smooth tapered antennae situated at anterior end ( Figure 10B, C View Figure 10 ). Pair of palps with massive palpophores and short round palpostyles. Two pairs of eyes arranged trapezoidally (anterior pair with space 1.3–1.5 times as wide as that of posterior pair); anterior pair reniform and smaller; posterior pair round and larger (subdermal eyes unclear in holotype and some other large specimens). Trapezoidal dark pigmentation with midlongitudinal white slit present on dorsal anterior surface of prostomium ( Figure 10C View Figure 10 ), though pigmentation unclear in type specimens probably because of discoloration from long-term preservation ( Figure 10B View Figure 10 ); dark pigmentation with white slits often present also on dorsal surface of palpophore ( Figure 10C View Figure 10 ).
Peristomium slightly longer than subsequent chaetigers, with four pairs of tentacular cirri of unequal length; posterior dorsal tentacular cirri longest, reaching back to chaetigers 2 to 9 (chaetiger 4 in holotype, usually chaetigers 3 to 4); anterior dorsal tentacular cirri next longest.
Proboscis with pair of brown jaws, each with 13 or 14 teeth in holotype. Black paragnaths with usually sharply pointed tip present on both maxillary and oral rings ( Figure 10 View Figure 10 B–E). Broad transverse band of many paragnaths (about 300 in holotype) present in dorsal and ventral areas of oral ring, covering groups V and VII–VIII, except for group VI remaining in compact clusters separated from other paragnaths; broad band of 8–10 irregular rows of paragnaths in dorsal and ventral areas, tapering to two or three rows in lateral sides ( Figure 10E View Figure 10 ); paragnaths larger in more distal row (on everted proboscis). Paragnath numbers on other groups in holotype (range for all examined specimens in parentheses) as follows, group I: 3 (1–4, n = 26) in longitudinal arrangement, II: 24 on each side in three arched rows, total 48 (28–58, n = 16), III: 9 (3–17, n = 29) in triangular or rhomboid patch, IV: 26 on right and 24 on left in three arched rows, total 50 (35–68, n = 14), VI: 12 on right and 11 on left in circular cluster, total 23 (15–27, n = 18).
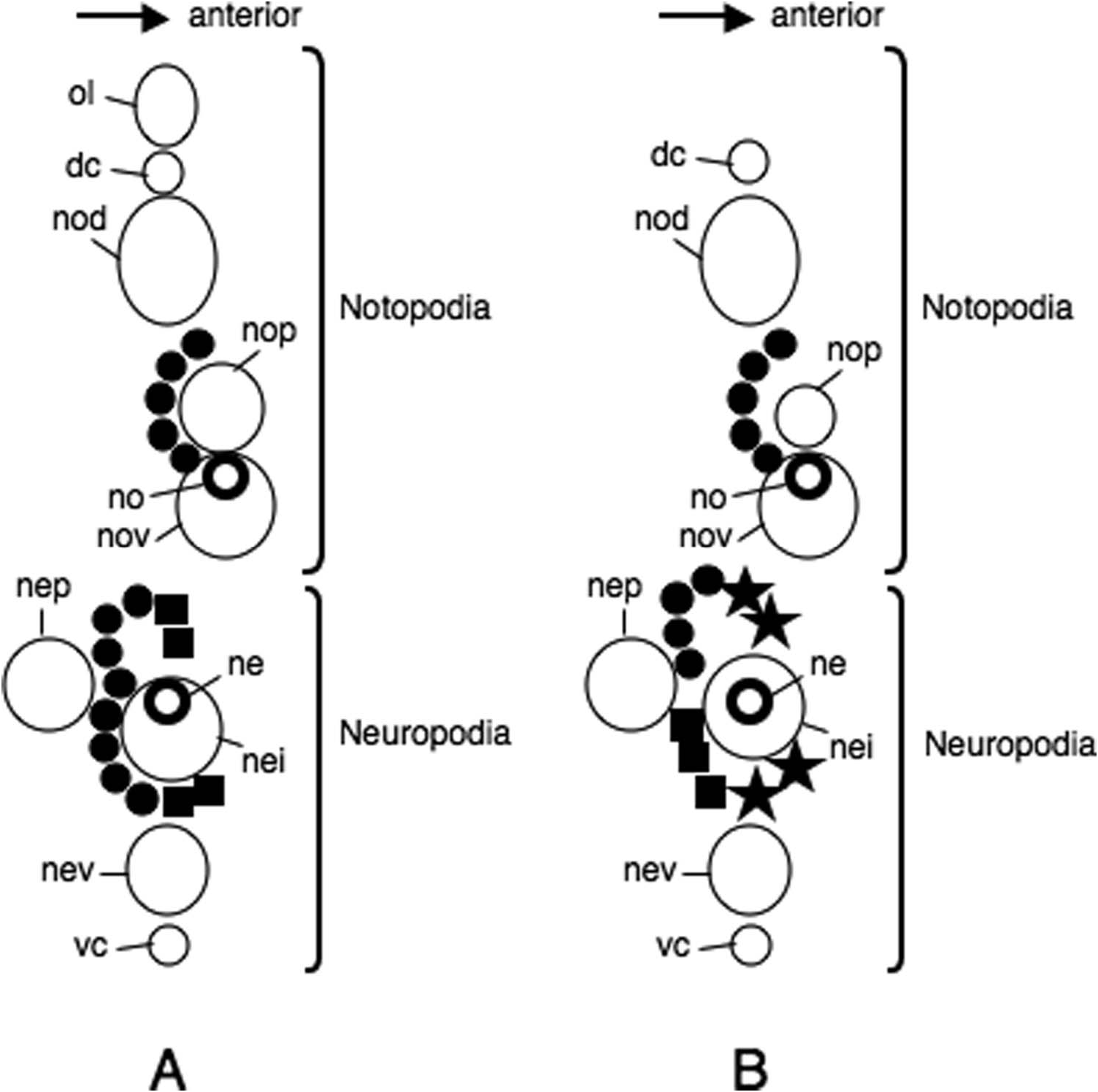
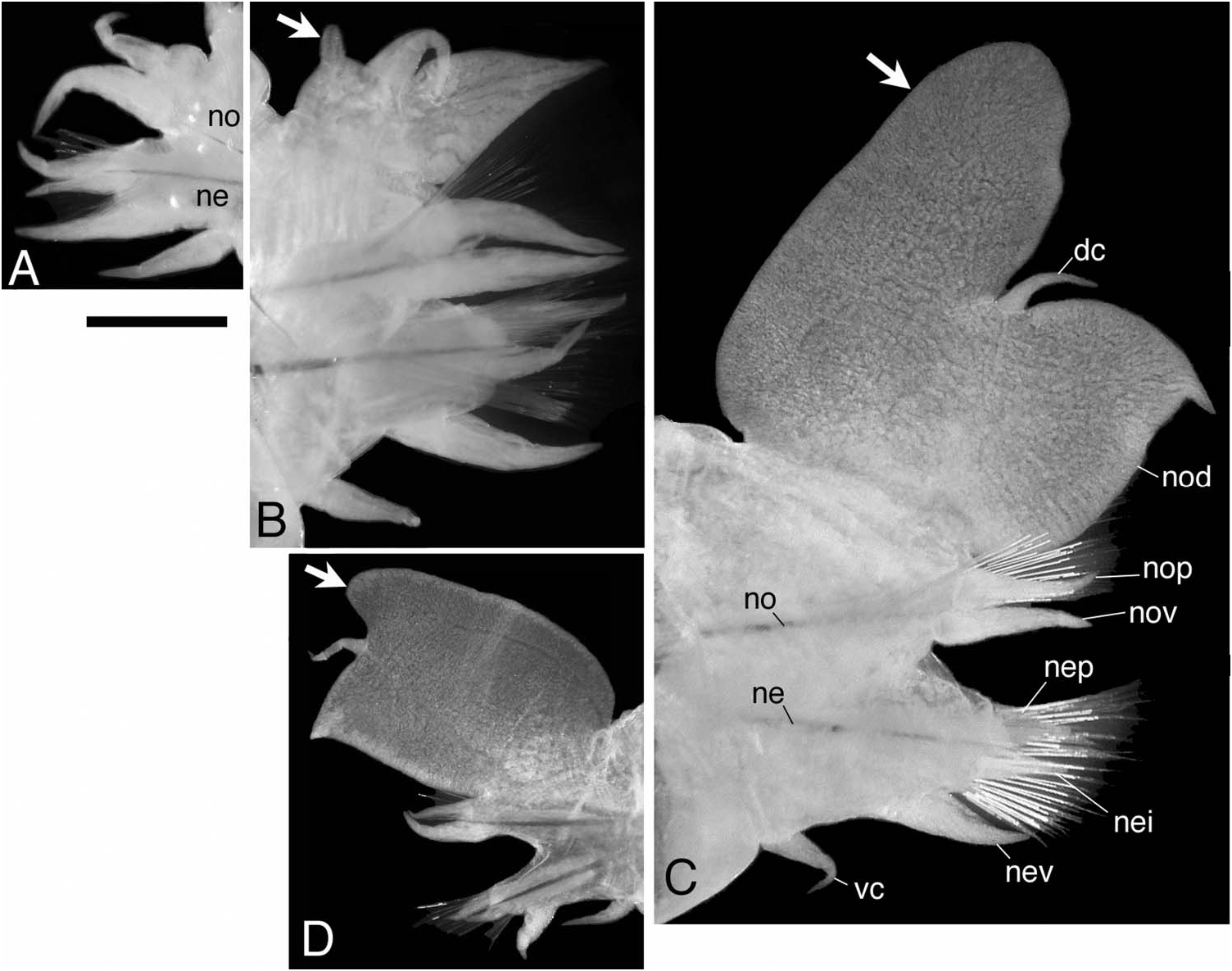
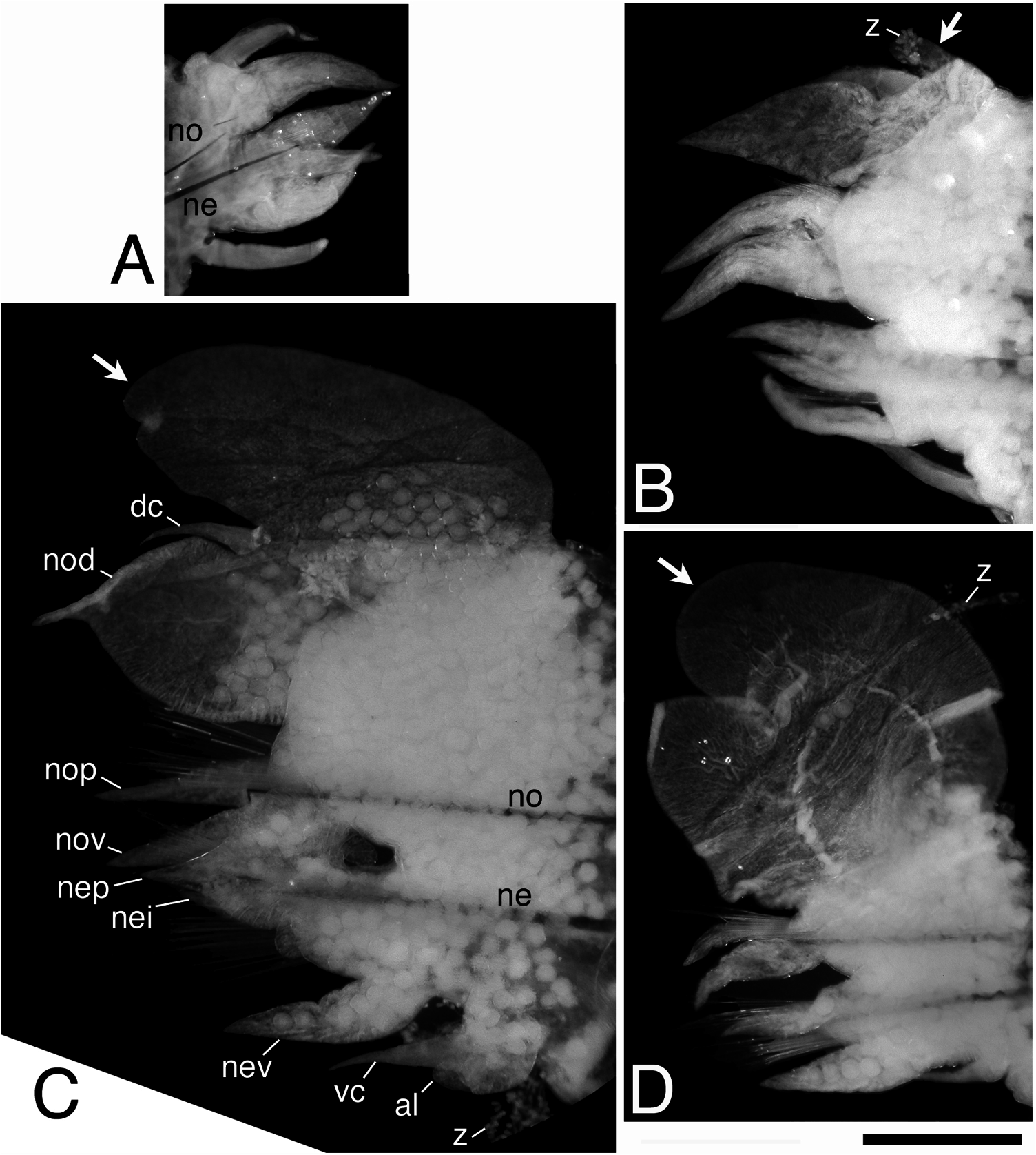
Parapodia of first two chaetigers sub-biramous, all following parapodia biramous. Sub-biramous parapodia with thin notoacicula and reduced notopodia consisting of dorsal cirrus and notopodial dorsal ligule, and with ordinary neuroacicula and ordinary neuropodia ( Figure 11A View Figure 11 ).
Notopodia consisting of dorsal cirrus, dorsal ligule, prechaetal lobe and ventral ligule in biramous parapodia. Notopodial dorsal ligule slender with tapering tip in anterior parapodia, expanded to large triangular with prominent ovoid lobe developing above dorsal cirrus in middle and posterior parapodia; first ovoid lobe above dorsal cirrus appearing at chaetigers 13–15 (chaetiger 13 in holotype, Figure 11B View Figure 11 ), except at around chaetiger 20 in some juveniles; notopodial dorsal ligule most enlarged in middle parapodia (chaetigers 50–100, Figure 11C View Figure 11 ), gradually diminishing in size in posterior parapodia ( Figure 11D View Figure 11 ). Notopodial prechaetal lobe and notopodial ventral ligule slender with tapering tip throughout; notopodial prechaetal lobe subequal to notopodial ventral ligule almost throughout. Dorsal cirri slender, tapering, reaching to tip of notopodial dorsal ligule in anterior parapodia, about half length to tip of notopodial dorsal ligule in middle and posterior parapodia.
Neuropodia consisting of inferior lobe, postchaetal lobe, ventral ligule and ventral cirrus throughout; all neuropodial lobes and ligule slender with tapering tip throughout, of similar length. Ventral cirrus slender with tapering tip ( Figure 11 View Figure 11 A–D).
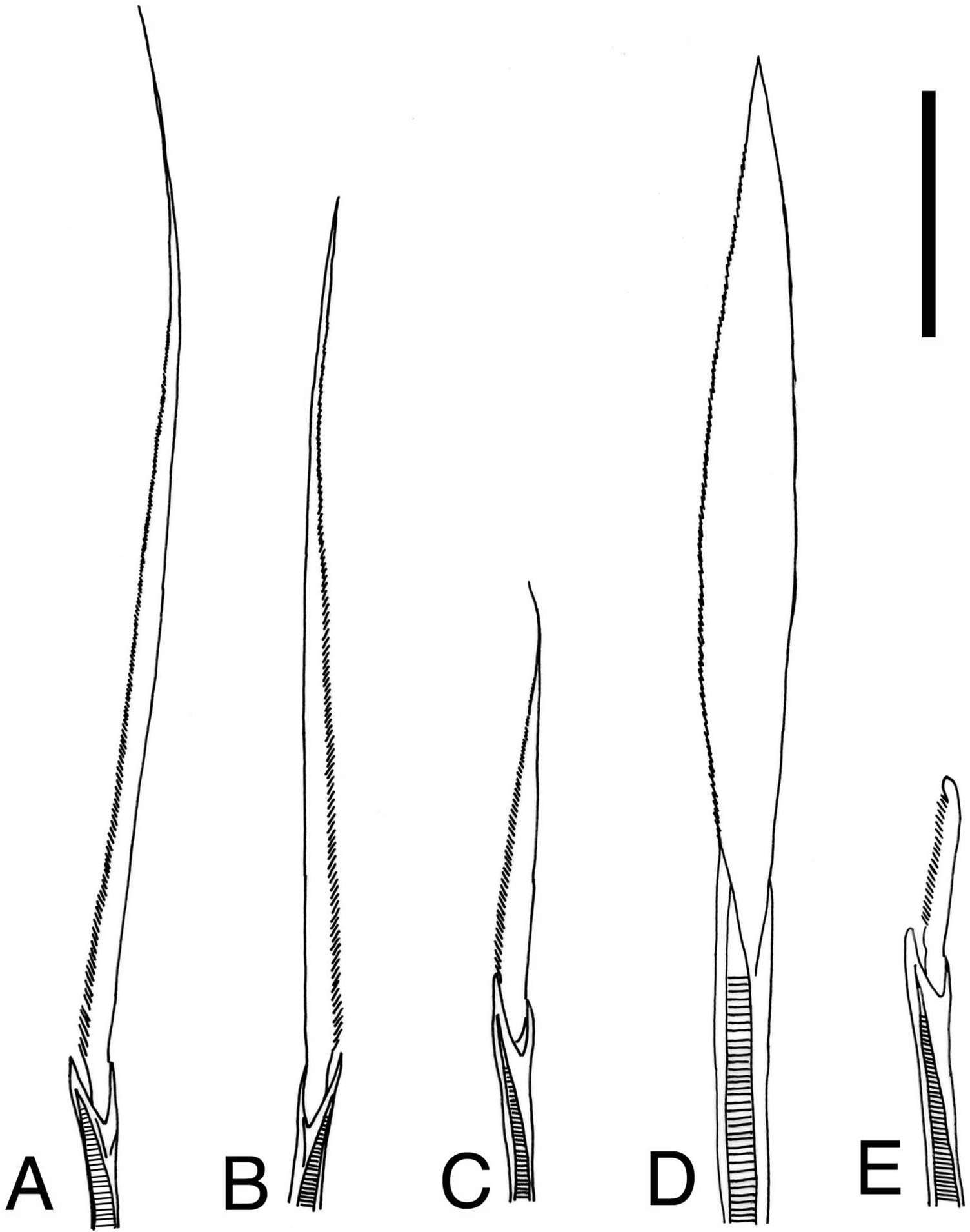
Notochaetae all homogomph or sesquigomph spinigers; blades long with finely serrated edge ( Figures 4A View Figure 4 , 12A View Figure 12 ). Numbers of notochaetae in holotype about 50 in parapodia 12 and 13, about 40 in parapodium 65, and about 10 in parapodium 125. Upper neurochaetae consisting of heterogomph spinigers with short serrated blades ( Figure 12C View Figure 12 ) at superior / anterior position (5 in parapodium 2, 15 in parapodium 12, 10 in parapodium 65, and 1 in parapodium 125 in holotype), and homogomph or sesquigomph spinigers with long serrated blades ( Figure 12B View Figure 12 ) at inferior / posterior position (13 in parapodium 2, 25 in parapodium 12, 20 in parapodium 65, and 10 in parapodium 125 in holotype) ( Figure 4A View Figure 4 ). Lower neurochaetae consisting of homogomph or sesquigomph spinigers with long serrated blades at superior / posterior position (25 in parapodium 2, 40 in parapodium 12, 20 in parapodium 65, and 7 in parapodium 125 in holotype), and heterogomph spinigers with short serrated blades at inferior / anterior position (15 in parapodium 2, 20 in parapodium 12, 15 in parapodium 65, and 1 in parapodium 125 in holotype). One or two heterogomph falcigers with slender serrated blades present in lower neurochatae throughout in small juveniles (less than 1.5 mm BW) ( Figure 12E View Figure 12 ).
Pygidium with anus on dorsal side, with pair of cylindrical slender anal cirri.
Non-type epitokes
One incomplete female 5.5 mm BW. Three almost complete males 70–160 mm BL, 2.5–5.0 mm BW, with 130–150 chaetigers. Eyes slightly enlarged at least in males (enlargement of eyes unclear in female) ( Figure 10F, G View Figure 10 ).
Parapodia and chaetae in anterior body (first 17–18 chaetigers in males, 33 chaetigers in female) similar to those of atokes, with slight changes in males; dorsal cirri of first seven chaetigers and ventral cirri of first five chaetigers thickened in males ( Figure 13A, B View Figure 13 ), with no marked change in female ( Figure 14A, B View Figure 14 ).
Parapodia and chaetae in middle body (70–90 segments in males, 40 segments in female) modified ( Figures 13C View Figure 13 , 14C View Figure 14 ). Additional lobes present at upper and lower portions on base of ventral cirri; upper lobe smaller, lower lobe larger, appearing from chaetigers 18–19 to chaetigers 87–104 in males, from chaetiger 34 to chaetiger 77 in female, becoming much larger in male than in female. Neuropodial postchaetal lobe developing into large round flat lamella with single tiny protrusion on lateral edge, and with digitate protrusion on inner side of lower edge in males; neuropodial postchaetal lobe expanded to triangular lamella with tapering tip in female. Notopodial ventral ligule expanded remarkably to triangular lamella in males, slightly enlarged in female. Dorsal cirri with serration on lower edge, slightly elongated in males; dorsal cirri with neither serration nor elongation in female. Ventral cirri with minute serration on upper edge, slightly elongated in males; ventral cirri with neither serration nor elongation in female. Atokous chaetae substituted by epitokous natatory ones ( Figure 12D View Figure 12 ) completely in males, and incompletely (several atokous spinigers remaining) in female; blade of epitokous natatory chaetae flat and wide, with minutely serrated edge on one side, and tapering tip.
Parapodia and chaetae in posterior body (about 50 segments behind chaetigers 87–104 in males, behind chaetiger 77 in female) ( Figures 13D View Figure 13 , 14D View Figure 14 ) similar to those of atokes. Many protozoan colonies attached to parapodia ( Figures 13D View Figure 13 , 14 View Figure 14 B–D).
Coelom of female filled with eggs (full-grown oocytes); eggs spherical or of somewhat irregular shape, 130–150 µm in diameter in fixed specimen.
Variation of paragnath number
Paragnath numbers in groups I–VI in Nectoneanthes uchiwa sp. nov. in three localities are summarized in Table 2 View Table 2 .
Reproduction
Only four epitokous specimens were examined, which were collected from Ariake Sea in Japan in spring. A female was collected around 23:00 (1 hour after night high tide in spring tide) on 20 April 2004. Three males were collected around 01:00 (1.5 hours after night high tide in spring tide) on 8 May 2004 ( Hanafiah et al. 2006). Epitokous males and females were also collected from China in April to May ( Wu et al. 1985).
Habitat
Muddy bottom in intertidal or shallow subtidal areas (up to 20 m deep) within estuarine embayments.
Geographical distribution
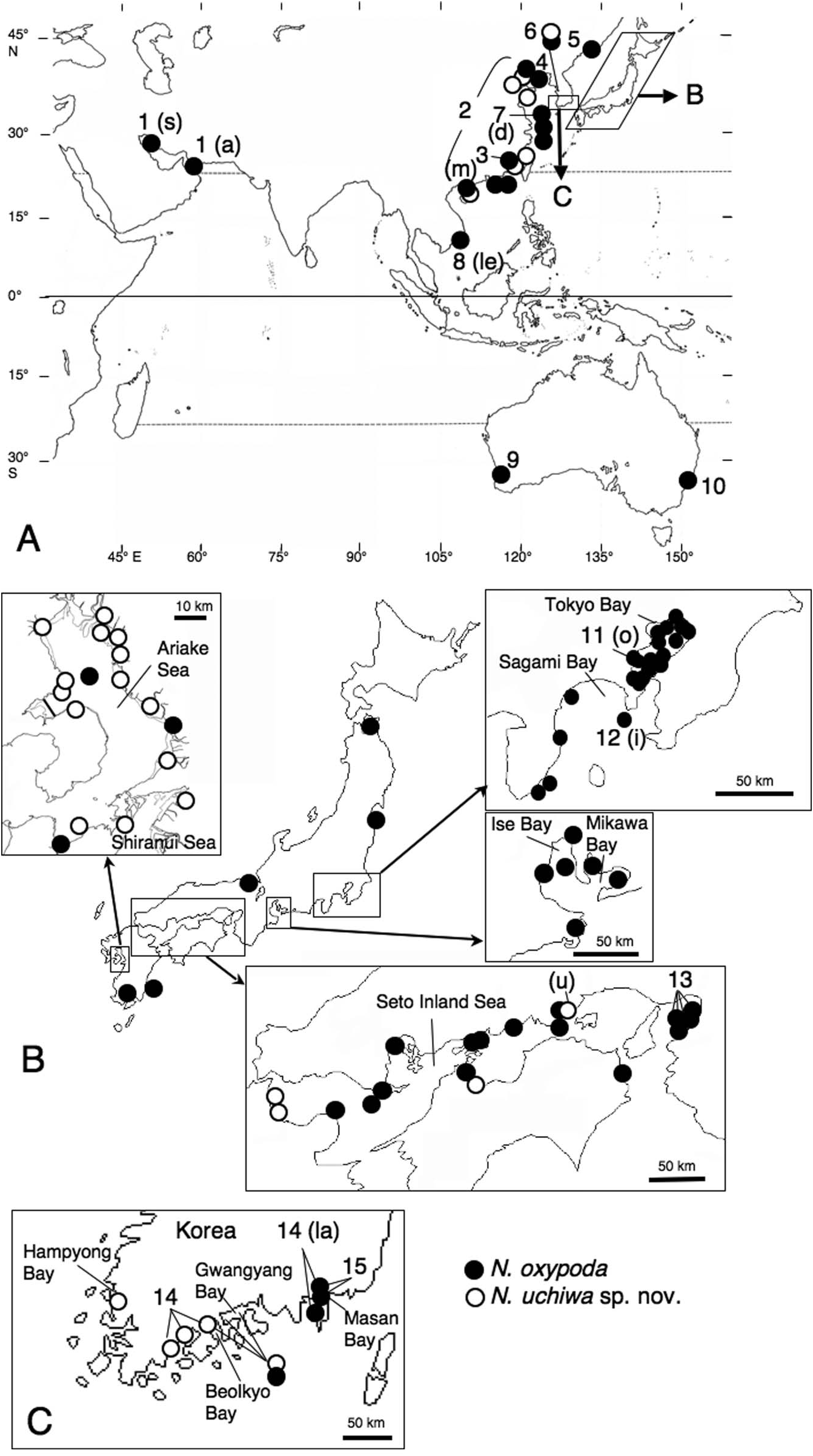
Western Japan (Seto Inland Sea, Ariake Sea, and Shiranui Sea), Korea, China ( Figure 9 View Figure 9 ).
Etymology
The specific name refers to a Japanese word (Uchiwa) meaning a paper hand fan used on hot summer days in Japan. The hand fan resembles the characteristic enlarged notopodial lobe of this species. The Japanese name of this species (Uchiwa-gokai) also refers to the same word.
Remarks
This species is easily distinguishable from N. oxypoda by the presence of a broad band of many paragnaths in both dorsal and ventral areas of the oral ring of the proboscis. This species also has larger numbers of paragnaths in three groups on the maxillary ring of the proboscis than N. oxypoda , though the ranges of their paragnath numbers overlap.
Heterogomph falcigers were found only in small juveniles (around 1.0 mm BW) collected in Korea, and not found in slightly larger juveniles (1.5 mm BW) collected in Japan. The morphological characteristics except for the presence of the juvenilespecific falcigers were adequately drawn in Okuda (1933), Imajima (1972, 1996), Paik (1972, 1973), Wu et al. (1985) and Khlebovich (1996) for the atokous form, and in Wu et al. (1985) for the epitokous form.
Worms of this species were used as bait for fishing in the Seto Inland Sea. According to Okuda (1933) and Mori et al. (1932), they were referred to as “ Katakogare ” or “ Hiiru ” by local people around Kojima Bay in Okayama Prefecture and as “ Umba ” or “ Shiomushi ” in Hiroshima Prefecture. According to Izuka (1912), they were referred to as “ Chibiiru ” around the Kojima Bay. Because both N. uchiwa sp. nov. and N. oxypoda inhabited the Kojima Bay as shown in the present study, the local people might distinguish these two species, naming them differently.
According to Wu et al. (1985), this species had important economic significance as an excellent bait for fishing in China, with a large amount of worms exported to Japan under the commercial name “Golden nereis”. Saito et al. (2011) also reported that this species is imported from China and sold in western Japan in winter under the commercial Japanese name “ Aka-kogane ”, meaning red gold. This is the reason why I designate old specimens of Izuka’s collection collected from the Kojima Bay in 1906 as the type series in the present study, despite the fact that the type locality was lost through coastal development (see Discussion). I am afraid that morphologically similar but genetically different populations of this species may be introduced to Japanese waters by recent commercial transportation. The Izuka’s collection, on which Izuka (1912) based the first description of this species under the name of “ Nereis oxypoda ”, has been safely preserved in good condition in NSMT. The drawing of the paragnath arrangement on a proboscis, which is the diagnostic characteristic of this species, was first provided by Okuda (1933) based on specimens collected from the Kojima Bay in April, 1933 by Prof. Tohru Uchida (Hokkaido Imperial University); I have not yet found the specimens.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Nectoneanthes uchiwa
| Sato, Masanori 2013 |
Nectoneanthes oxypoda sensu Imajima, 1972
| Saito H & Niwa N & Kawai K & Imabayashi H 2011: 47 |
| Hanafiah Z & Sato M & Nakashima H & Tosuji H 2006: 209 |
Neanthes
| Wilson RS 1988: 7 |
Neanthes (Nectoneanthes) oxypoda
| Wilson RS 1984: 220 |
Nectoneanthes oxypoda: Imajima, 1972: 113–118
| Yamanishi R & Sato M 2007: 183 |
| Lee J-H & Park J-Y & Lee H-G & Park H-G & Kim D 2003: 189 |
| Imajima M 1996: 144 |
| Khlebovich VV 1996: 115 |
| Yang D & Sun R 1988: 39 |
| Wu BL & Sun R & Yang D 1985: 164 |
| Paik E-I 1973: 82 |
| Imajima M 1972: 118 |
| Paik E-I 1972: 135 |
Neanthes oxypoda: Imajima and Hartman, 1964: 145
| Imajima M & Hartman, O 1964: 145 |
Nereis (Neanthes) oxypoda: Khlebovich, 1963: 55
| Khlebovich VV 1963: 55 |
Nereis oxypoda: Izuka, 1912: 171–173
| Monro CCA 1934: 362 |
| Okuda S 1933: 247 |
| Mori T & Tamura M & Makino K 1932: 3 |
| Izuka A 1912: 173 |