
Brachypeplus glaber LeConte, 1878: 398
|
publication ID |
https://doi.org/10.11646/zootaxa.3734.2.9 |
|
publication LSID |
lsid:zoobank.org:pub:06352BA1-0C93-454B-A691-D7A62835E298 |
|
DOI |
https://doi.org/10.5281/zenodo.6148839 |
|
persistent identifier |
https://treatment.plazi.org/id/03A887F1-2C41-7C24-A8D2-FEFB9FE27B02 |
|
treatment provided by |
Plazi |
|
scientific name |
Brachypeplus glaber LeConte, 1878: 398 |
| status |
|
Brachypeplus glaber LeConte, 1878: 398
Type material examined. Holotype [MCZ] ♂—pecimen and label images available at: http:// insects.oeb.harvard.edu/mcz/FMPro?-DB=Image.fm&-Lay=web&-Format=images.htm&Species_ID=6961&-Find (Perkins 2010).
Other material examined. More than 100 non-type specimens were studied.
Adult diagnosis. Brachypeplus glaber differs from other members of the genus by the following combination of characters: overall bicolored appearance of the body with abdomen and thoracic ventrites piceous in color and rest of body reddish brown; reduced elytral and pronotal setation; prothorax densely punctate with scale-like reticulate microsculpturing between punctures; temples prominent and moderately protruding; serial punctures on elytra densely distributed and flat; and scutellum pentagonal.
Adult description. Parsons (1943) provided an updated revision of LeConte’s (1878) original description. Although rudimentary line drawings of the mouthparts and male genitalia were provided, little detail on species level characters was given and female genitalia were not described. The following is a more complete and modern description of the species.
Overall body shape elongate and mostly parallel-sided. Average length = 3.7 mm (n = 10), avg. width 0.9 mm (n = 10). Overall, body elongate, ~4× as long as wide, and exceedingly flattened. Body distinctly bicolored with abdomen and thoracic ventrites dark brown to piceous and rest of body reddish-brown (see Cline & Skelley 2013, Fig. 14). Surface sculpturing on dorsum distinct and consisting of “scale-like” circular microreticulations between punctures on the head, pronotum, and elytra. Body setation minute and only apparent at high magnification.
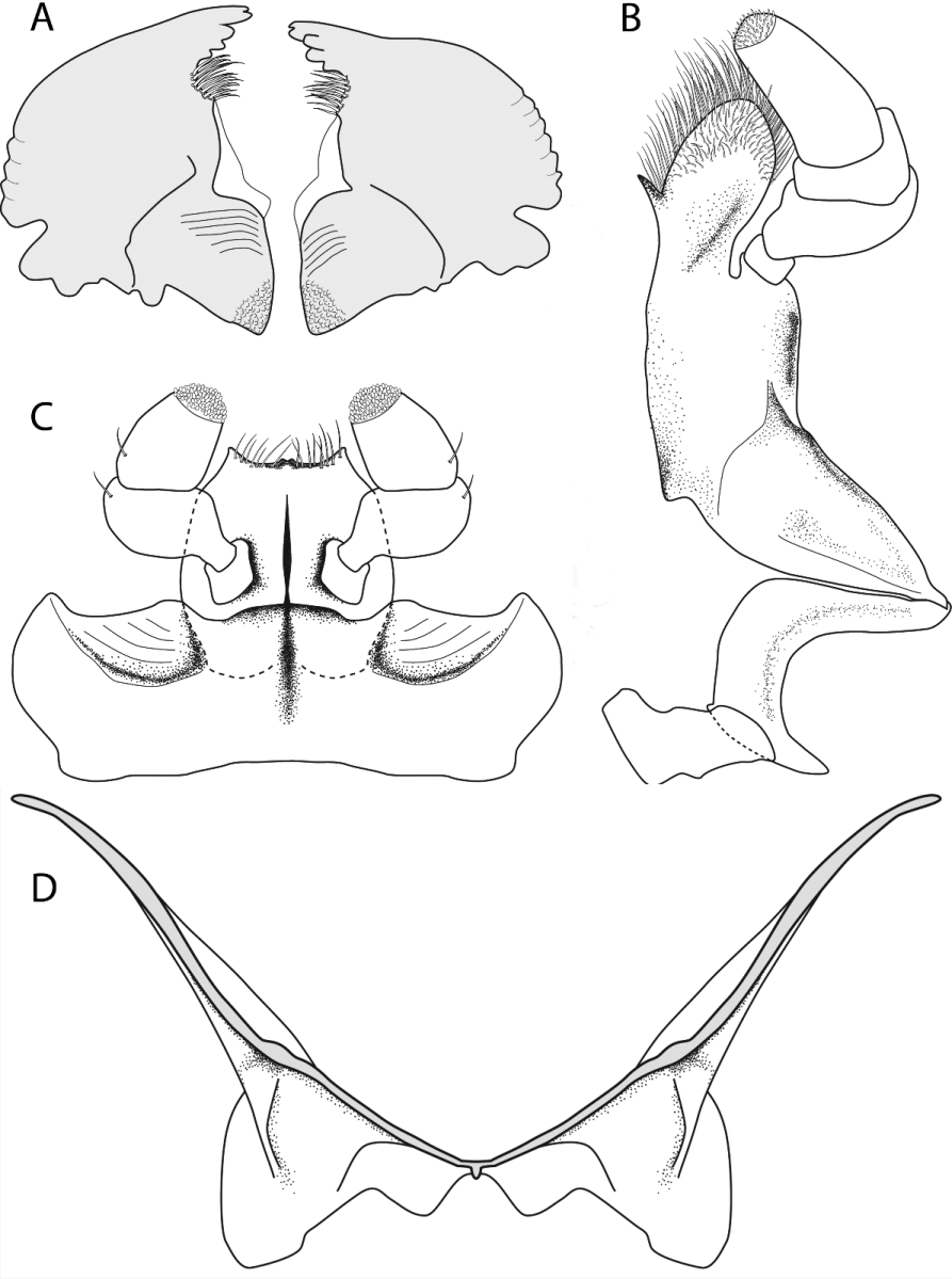
Head transverse (W:L = 1.00:1.55), with a broad transverse depression across the anterior part of the vertex. Dorsal postocular line extending posteriorly onto temple region, line faint but visible. Surface punctures circular, regular, slightly larger than eye facet with minute puncture often found near anterior margin of larger puncture but no setae visible. Interspaces between punctures microreticulate and shining, appearing scale-like, and 1–2 puncture diameters apart. Antennal grooves present, moderately impressed, and strongly convergent. Vertex flattened to shallowly concave across middle. Temples present, convergent, and slightly produced beyond outer eye margin. Eyes not protruding, finely faceted, interfacetal setae absent. Antennal club oval; club ~0.40× length of segments 1–8 combined. Terminal antennomere shorter than previous two segments combined; apex of terminal segment acuminate and with annulate apical region. Scape enlarged, broadly convex anteriorly, and ~2× longer than segment 2. Pedicel as long as wide, somewhat barrel-shaped to quadrate. Antennal segment 3 elongate, ~1.5× as long as segment 2; segments 4–7 similar in size; segment 8 somewhat discoidal. Labrum distinct, but short, anterior margin weakly, indistinctly bilobed. Mandibles with apex bifid; 2–3 subapical teeth present distal to prosthecal region; prosthecal region with large apical field of elongate setae; molar region large with broadly defined grinding region with oblique rows of grooves; lateral margin of mandible slightly scalloped ( Fig. 1 View FIGURE 1 A). Lacinia of maxilla with moderately narrowed lobe, apical third of lobe with border possessing elongate setae; large spinose process present along medial border, separating setae from rest of lobe ( Fig. 1 View FIGURE 1 B). Maxillary palpi with terminal segment conical and equal to segments 1–3 combined. Labial palpi with terminal segment barrel-shaped and subequal to segments 1–2 combined ( Fig. 1 View FIGURE 1 C). Mentum highly transverse with antero-lateral angle produced and acuminate, medial angle not prominent along anterior margin, posterior margin straight to slightly concave.
Pronotum wider than long (W:L = 1.5:1.0), appearing somewhat transverse, sides gently sloping to slightly deflexed lateral margin. Anterior and posterior angles rounded. Lateral margins narrowly explanate, more explanate in the lateral posterior regions; lateral edge slightly convex medially and gradually sloping to anterior and posterior angles. Anterior and posterior margins slightly lobed medially. Surface punctures similar to those on head, except minute punctures bearing short pale gray setae, only visible under high power (100× or greater). Prothorax with space between anterior margin of procoxae and anterior margin of prosternum ~3× longer than space between posterior margin of procoxae and posterior margin of prosternum; some widely scattered punctures and transverse lines present. Prosternal process narrow between procoxae and greatly expanded posterior to procoxae.
Mesonotum with scutellum pentagonal; surface punctation and sculpturing similar to that of head, no setae present. Mesoventrite at same level as metaventrite, mostly flattened, with disc smooth and shining, some diagonal lines apparent. No clear delimitation of mesoventrite from mesepimeron. Mesoventrite meeting metaventrite in truncate junction.
Metaventrite wider than long (W:L = 1.45:1.00); metaventral disc surface punctation and sculpturing similar to that of head. Postcoxal lines of mesocoxae not significantly deviating from coxal cavity, with no apparent axillary space present. Posterior margin of metaventrite with shallow triangular notch present medially. Metanepisternum narrow, and tapering to half its width at posterior fifth of structure, with posterior apex truncate. Metendosternite moderately sclerotized, exceedingly wide and broadly flanged; lateral arms of furca widely divergent and thin; lateral portion of ventral process weakly developed and thin, obliquely attaching to lateral lobe of furcal stalk; stalk of furca broad with two large lateral lobes and two smaller medial lobes; ventral median flange of furca greatly reduced to small area at midline ( Fig. 1 View FIGURE 1 D).
Elytra greatly reduced, exposing last 3 visible abdominal tergites; individually each elytron longer than wide (L:W = 2.1:1.0); lateral margins faintly explanate with explanation continuing anteriorly around humeri and onto anterior margin, but not extending posteriorly to postero-lateral angle; apices separately rounded; pubescence short, golden, and only present in lateral region; surface with alternating rows of large and small punctures, all punctures faintly impressed; surface sculpturing scale-like as on rest of body, shining. Humeral angles moderately prominent.
Abdominal ventrite 1 projecting between metacoxae with broadly rounded anterior projection. Abdominal ventrites 1 and 2 similar in length, and 3 and 4 similar in length with 3 or 4 approximately 1.7 times as long as either 1 or 2. Hypopygidium 1.5× as long as visible ventrite 4, and with posterior margin arcuate in females, and more truncate in males. Pygidium with posterior margin indentate in females and truncate to broadly arcuate in males.
Legs normal with no apparent sexual dimorphism. Femoral shape typical of most nitidulids with widest point near middle, only distal half of femur canaliculate for reception of tibiae. Tibiae with unequal apical spurs. Protibial shape evenly curved along outer margin, with lateral edge crenulate; apical margin bearing short teeth. Meso- and metatibial shape and armature similar to that of protibiae, but with more and longer apical teeth present. Tarsomeres 1–3 expanded, appearing almost lobed, with dense setose pads present on ventral surface, tarsomeres 4–5 simple with no expansion or setal pads; claws simple with no teeth present.
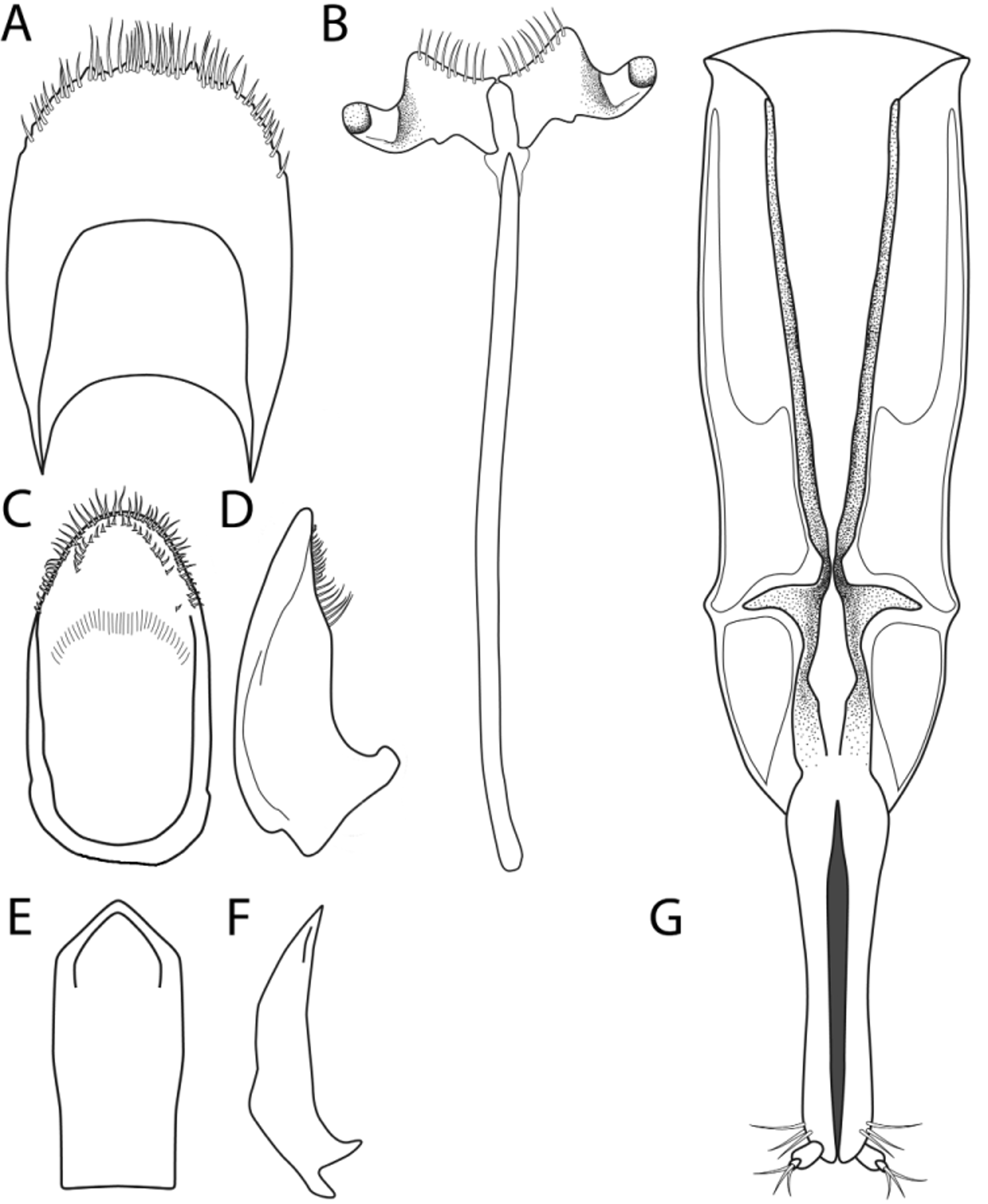
Male genitalia well sclerotized. Anal sclerite broadly rounded at apex with small tubercles present along margin, apical tuft of setae prominent with setae becoming shorter laterally ( Fig. 2 View FIGURE 2 A). Spiculum gastrale broadly flanged with relatively few primary setae present along apical margin, typically less than 15 ( Fig. 2 View FIGURE 2 B). Tegmen with apical region bearing a subapical setose ridge; apex densely setose ( Fig. 2 View FIGURE 2 C); in lateral view with setae more elongate basally and becoming shorter apically ( Fig. 2 View FIGURE 2 D). Median lobe of aedeagus with apex acutely rounded, apical lateral angles defined, base somewhat constricted, and apical opening simple ( Fig. 2 View FIGURE 2 E); in lateral view with distinct notch on dorsal margin approximately halfway down the length of the structure ( Fig. 2 View FIGURE 2 F). Female genitalia ( Fig. 2 View FIGURE 2 G) moderately well-sclerotized. Gonocoxites elongate, fused basally, base with diamond-shaped opening in middle, three subapical primary setae present apicolaterally just proximal to apical styli; apical styli welldeveloped with three primary setae originating from small apical pit. Valvifers elongate, widest near middle with slight constriction near basal third of structure.
Variation. Teneral specimens are not distinctly bicolored. Complete sclerotization and melanization takes 4–6 days to occur following adult eclosion (Habeck, unpub. notes). Males and females exhibit three main external morphological differences: 1) posterior margin of hypopygidium is more rounded in females and more truncate in males; 2) posterior margin of pygidium is indentate in females and truncate to broadly rounded in males; and 3) males typically have a so-called “anal sclerite”, which is part of the male genital capsule, protruding from between the pygidium and hypopygidium.
Geographic distribution. Brachypeplus glaber was previously known from several sites in Florida (Dunedin, Capron, Lake Worth, St. Augustine, and Enterprise), and one site in southeastern Georgia (St. Simon’s Island) (LeConte 1878; Hamilton 1894; Parsons 1943). We note this species herein from several other localities in Florida and Georgia, as well as coastal Alabama, Mississippi, South Carolina, and North Carolina. The updated distribution of the species coincides with a rather widespread occurrence throughout much of the coastal areas of the southeastern United States. Brachypeplus glaber is likely widely distributed throughout the known range of its host plant Sabal palmetto from extreme southern Virginia southward around peninsular Florida, and westward along the Gulf Coastal states into Louisiana or perhaps eastern Texas. However, correspondence with a prominent beetle collector and specialist suggests that B. glaber likely does not occur in eastern Texas (Ed Riley, pers. comm.), as many attempts to collect beetles within and among palms has yielded no specimens of this species. The ornamental palm industry has transplanted specimens of cabbage palms into many regions throughout the southeastern United States that possess an amenable climate. Therefore, there is potential for a much more extensive distribution; however, to date there have been no collection events of this beetle other than from the likely native range of the host plant. A companion paper on the biology, ecology, natural history, and biogeography of this species is currently being completed (Cline et al., unpub. manuscript).
Etymology. The specific epithet is apparently indicative of the highly reduced body setation of the adult form. Some Brachypeplus species have densely distributed setae, and B. glaber is rare in having highly reduced body setation.
Egg description. Average length of an egg is 1.04 mm (n = 11, range = 0.99–1.11 mm). The average width of an egg is 0.18 mm (n = 8, range 0.13–0.24 mm). Overall, cigar-shaped with a surface sculpturing pattern of a reticulated polygonal type, translucent to opaque cream colored.
Third instar larval diagnosis. Brachypeplus glaber differs from other known members of the genus by the following combination of characters: entire surface rugose with fine bumps and tubercles on all body segments; each thoracic and abdominal tergite with well-developed and defined plates on each side of the midline; terminal antennal appendage less than half the length of terminal antennomere; mesal surface of mandible anterior to mola and posterior of prostheca with 3–4 well-developed, blunt, heavily sclerotized teeth; metacoxal cavities broadly separated by ~5× the width of a coxal cavity; anal shield with prominent but small pregomphi oriented in an oblique dorsal-posterior manner; anal shield with three pairs of terminal urogomphal processes, one lateral and two posterior.
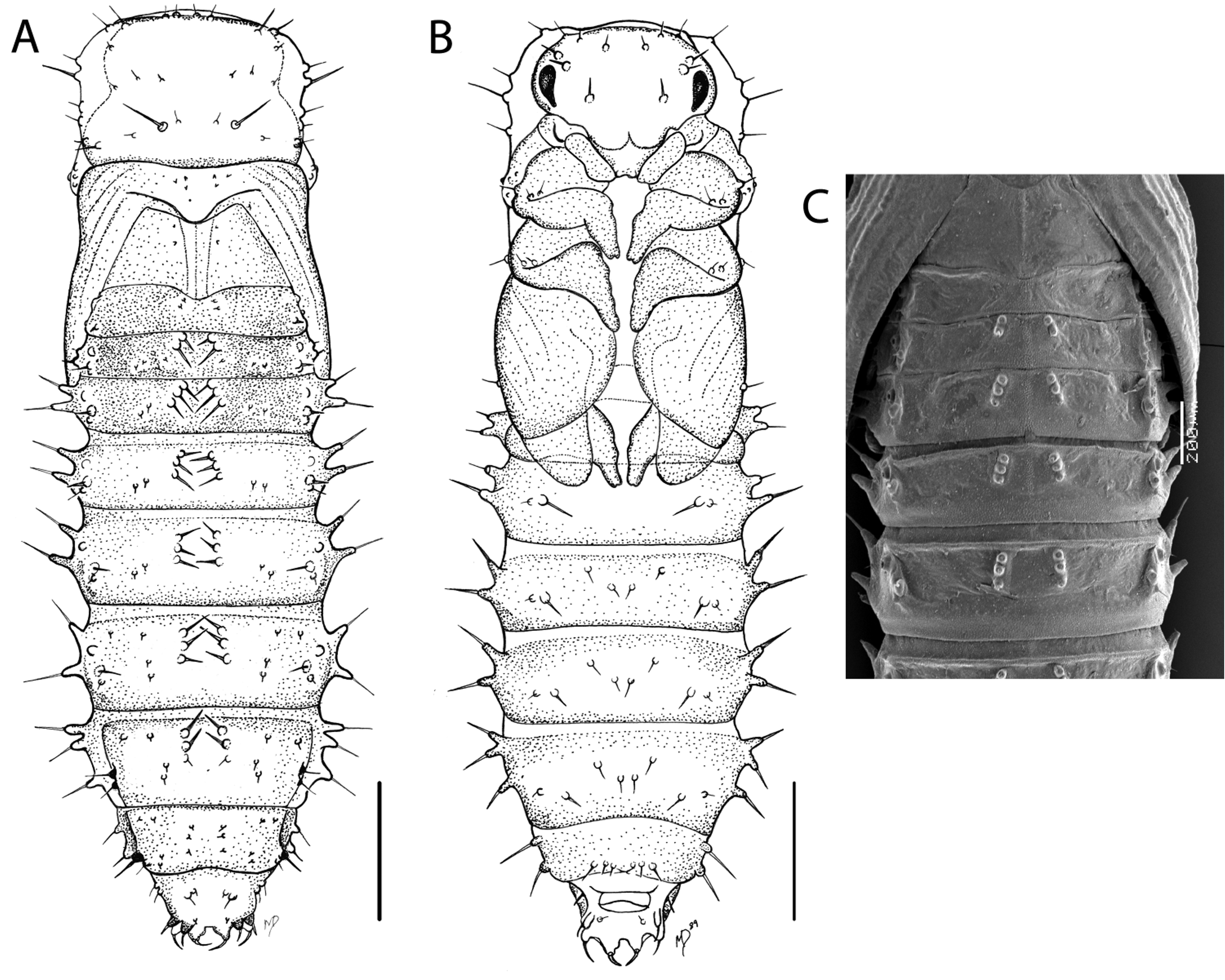
Third instar larval description. Body elongate,>4× as long as wide, sub-parallel, moderately dorso-ventrally flattened, widest near A4–A5. Average length 4.7 mm (n = 4), average width 1.1 mm (n = 4). Urogomphi and pregomphi present on distinct anal shield of A9 tergite. Body typically cream colored to light tan. Head well sclerotized. Legs and antennae clearly visible and distinct. All thoracic and abdominal tergites with paired lightly sclerotized oval to circular plates bearing setiferous tubercles. Abdominal segments 1–7 with sharply angulate lateral margins near middle. Dorsal body setation scarce, mostly limited to setiferous tubercles or sparsely distributed on surface. All spiracular openings located apically on short tubes.
Head prognathous, slightly narrower than T1, surface with few scattered short setae. Three prominent setae are located along the lateral margin, and two prominent setae are located on the vertex on each side of the head (Fig. 3A). Posterior dorsal margin of head capsule broadly emarginate, not deeply indentate. Frontal sutures separate at base with no epicranial stem present. Vertex slightly concave medially. Stemmata inconspicuous. Antennae 3- segmented; segment 2 twice as long as segment 1, with sensorium present at apex that is half the length of segment 3; segment 3 elongate with three prominent apical setae, two of which are oriented obliquely and one extending straight from apex. Frontoclypeal suture indistinct. Clypeal region with one prominent seta located near midpoint along lateral margin. Clypeolabral suture well defined. Labrum with anterior margin shallowly emarginate, two pairs of prominent setae present, one pair located at each lateral corner and the other near the midline. Mandibles heavily sclerotized (Fig. 3B); two prominent setae located along lateral margin; apex acute with multidentate dorso-medial edge, prostheca complex with multi-fringed and spinose processes; mola with prominent transverse and oblique bands of asperities and a thickened, lobed anterior grinding surface.
Pronotum with dorsal plate larger than any other thoracic or abdominal tergal plate; longer, narrower, and more quadrate than subsequent thoracic tergites; setiferous tubercles oriented along anterior, lateral, and posterior margin of plate. Meso- and metathoracic terga with plates more oval and smaller than those on prothoracic terga, and bearing fewer setiferous tubercles. Abdominal tergites 1–7 with lateral margins angulate, with prominent lateral setae extending from edge of angle. All abdominal tergites with paired oval to circular lightly sclerotized plates bearing setiferous tubercles (see Fig. 3C for close-up of A7 plate). Abdominal segments 4 and 5 wider than other abdominal or thoracic segments. Abdominal tergite 8 without markedly laterally angulate margin, lateral angle not prominent and occurring posterior to midpoint of segment; thoracic tergite with sclerotized plate appearing quadrate. Abdominal tergite 9 with prominent and heavily sclerotized anal shield; segment 9 subequal in length to segment 8; no lateral angulation present. Anal shield with well-defined scalloped anterior margin when viewed from above (Fig. 3E); numerous basal, lateral, and apical setae present, all oriented laterally, obliquely or posteriorly; pregomphi and urogomphi present and not fused to one another. Pregomphi and urogomphi when viewed laterally (Fig. 3D) with sharply acute apices and associated setae; pregomphi slightly less prominent than urogomphi.
FIGURE 3. Mature 3rd instar larva of B. glaber . A) split dorsal (left) and ventral (right), habitus line drawings; B) left mandible, line drawing, dorsal view; C) abdominal tergite 7; D) anal shield of abdominal tergite 9, line drawing, lateral view; E) anal shield of abdominal tergite 9, line drawing, dorsal view, scale bar = 0.1 mm; F) SEM of head, dorsal view; G) SEM of abdominal tergites, dorsal view; H) SEM of anal shield, oblique view.
Pupal diagnosis. Pupae of this species can be distinguished by the following combination of characters: setiferous tubercles along midline of tergites 5–7 arranged in semi-circular manner and setiferous tubercles on tergites 2–4 oriented in an oblique manner; tergite 9 with three minute non-setiferous tubercles along midline and four apical and two subapical large setiferous tubercles; setae always longer than associated tubercle on the lateral tubercles on all ventrites.
Pupal description. Body elongate, 3.6 mm long on average (n = 4) ( Figs. 4 View FIGURE 4 A, 4B). Orange-brown to yellowbrown in color. Head prognathous, almost completely deflexed under pronotum, strongly appressed to body, partially visible from above. One pair of setiferous tubercles near midline on vertex, one pair of setiferous tubercles near medial margin of each eye, and two pairs of setiferous tubercles (superior tubercle 3× larger than inferior tubercle) near frontoclypeal region of head. Clypeus strongly indentate medially. Mandibles, eyes, and antennae all readily apparent. Antennal apices with numerous conical protuberances when viewed under high power (100×).
Pronotum large and prominent, almost completely obscuring head in dorsal view, only small edge of head protruding beyond anterior margin of pronotum. Anterior margin nearly straight; one pair of setae present on anterior margin at each side of midline near middle of pronotum; a single submarginal seta present near lateral anterior angles; one marginal seta at lateral anterior angle. Lateral margin with 6 marginal setae present, some only visible under high power (100×), including one at the posterior angle. Posterior margin lacking marginal setae. Mesonotum with 4–6 scattered setae present on the surface. Mesothoracic wings extending beyond middle of 3rd abdominal tergite in dorsal view. Metanotum with two minute setae present near midline, only visible under high power (100×); posterior margin distinctly lobed at middle. Metathoracic wings extending beyond apex of mesothoracic wings; both wings curled ventrally beneath body. Metathoracic wings typically only partially visible at level of 2nd and 3rd abdominal segments, and extending to posterior margin of 3rd abdominal sternite.
Pro- and mesothoracic legs not obscured by wings; metathoracic leg segments mostly obscured by meso- and metathoracic wings, only tarsi clearly visible. Pro- and mesofemora with two setiferous tubercles located in apical third of leg segment on anterior face near tibial junction; metafemora with a single setiferous tubercle in apical third of leg segment on anterior face near tibial junction. All femora with a large setiferous tubercle on the lateral margin near the tibial junction.
Abdominal tergite 1 without medial setiferous tubercles ( Fig. 4 View FIGURE 4 C); one lateral setiferous tubercle present near lateral margin, seta minute and only visible under high power (100×). Tergites 2–8 with more well-developed lateral setiferous tubercles. Tergite 2 with two large setiferous tubercles on either side of the midline and oriented in an angular manner with the anterior tubercle closest to the midline. Tergites 3–4 with three large setiferous tubercles on either side of the midline and oriented in a similar angular manner as tergite 2; one pair of minute setae present between the large setiferous tubercles and the lateral margin. Tergites 5–7 with three large setiferous tubercles on either side of the midline oriented in a more semi-circular manner than those on tergites 3–4; one pair of minute setae present between the large setiferous tubercles and the lateral margin. Tergite 8 with three minute tubercles on either side of the midline oriented in a semi-circular manner as on tergites 5–7, each tubercle less than one-third the diameter of those on tergite 5–7 and with no associated seta. Tergite 9 with one small setiferous tubercle on either side of the midline, each tubercle slightly less than the diameter of those on tergites 5–7. Tergite 9 with four apical and two sub-apical large setiferous tubercles in posterior region.
Abdominal ventrites 1 and 2 obscured by metathoracic legs and meso- and metathoracic wings ( Fig. 4 View FIGURE 4 B). First abdominal ventrite smaller than all other ventrites, ~0.75 length of ventrite 2. Ventrites 1 and 2 lacking sublateral setiferous tubercles. Ventrite 1 with only one small setiferous tubercle along lateral margin; tubercle <0.5 length of posterior lateral tubercle on ventrite 2 and similar in size to anterior lateral tubercle. Ventrites 2–7 with paired lateral setiferous tubercles, anterior tubercle directed anteriorly on segments 2–6 and directed posteriorly on segment 7. Seta always longer than associated lateral tubercle on all ventrites. Ventrite 3 with paired sublateral setiferous tubercles between midline and lateral margin; medial tubercle in each pair larger than lateral tubercle. Ventrite 4 with lateral and sublateral setiferous tubercles as well as two additional pairs of tubercles near the midline in a U-shaped orientation, a third pair of minute setiferous tubercles is only visible under high power magnification (100× or greater). Ventrites 5 and 6 with similar arrangement of setiferous tubercles as ventrite 4. Ventrite 6 with posterior margin convex medially. Ventrite 7 with a pair of setiferous tubercles present on each side of the midline near the posterior margin. Spiracles present on abdominal segments 1–8; prominent on 1–6, and inconspicuous on 7–8.
Phylogenetic characters/character states from descriptions
Brachypeplus species are distributed worldwide, being mostly found in subtropical and tropical zones. The most closely related species to B. glaber , i.e., B. habecki Cline and Skelley (Cline & Skelley 2013) , is known only from Sabal mexicana Martius stands in extreme southern Texas, USA. Although this palm species is known to have a range extending southward into Mexico and Central America, the beetle species has only been recorded in Texas from a single specimen collected in a flight intercept trap. No one has looked within S. mexicana flower stalks to determine if more B. habecki are present there. Together, B. glaber and B. habecki form a clear species group based on shared external features (i.e., diminutive size, surface sculpturing, and tibial armature) of the adults, and are likely related to the Central American B. staphylinoides Sharp based on the conserved microsculpturing of the pronotum and appendages, which is not evident on other New World species. Microsculpture has been found to provide phylogenetic signal in other Nitidulidae in species-level phylogenetic reconstructions (Cline 2005). The use of the metendosternite to disentangle higher-level phylogenies at the generic and tribal levels has not been undertaken for Nitidulidae . Our results indicate that the adult metendosternite of B. glaber demonstrates clear similarities and differences from other Cillaeinae genera as well as other Nitidulinae genera. For example, the presence of two large lateral lobes on each side of the furcal stalk is shared with several other Cillaeinae genera, as well as the ventral median flange. Assessment of metendosternite characters of the adults in Nitidulidae has never been attempted and our data provide some insights into the usefulness of this character system. Authors Cline and Audisio are preparing a large dataset combining adult, larval, pupal, and molecular sequence data to ultimately provide an internal classification system for all Nitidulidae subfamilies, tribes, and genera. Metendosternite characters will be used in that analysis.
Characters from immature forms of Nitidulidae have never been utilized in any phylogenetic analysis at any hierarchical level; however, recent exploratory research was completed on several Meligethinae genera from Europe, Africa, and Asia (Reibaldi 2013; Audisio et al., unpubl.). Numerous character systems in larval Nitidulidae are ripe for investigation. This research demonstrated the usefulness of multiple characters for species- and generic-level assessments, as well as other characters that will be used in an upcoming treatment of world Nitidulidae larvae by the authors Cline and Audisio. At the species level within Brachypeplus , the number and placement of primary setae along the lateral margin of the mandible were found to vary between different, yet closely related, taxa, but remain stable within a species. The shape and setation of the pregomphi, not the urogomphi, also exhibited variability among species. The reverse is true for generic-level relationships with shape and placement of the urogomphi within Cillaeinae genera differing significantly. Likewise, the presence/absence of an apically oriented ventral tooth on the mandible varies between genera, but is conserved within a genus of Cillaeinae . Within Brachypeplus , the development of the spiracular tubes varies considerably with B. glaber having only moderately projecting lateral tubes with spiracular openings at the apex, and B. rubidus which has exceedingly long spiracular tubes with openings near the base of each tube. However, the length of spiracular tubes is typically associated with larval lifestyle within rotten, semi-liquid vegetal material and functions to ensure better respiration by projecting air passage entrances away from anaerobic fermenting substrates. The presence/absence of distinct sclerotized patches on the thoracic and abdominal tergites also varies between genera of Cillaeinae , e.g., they are absent in Colopterus and Macrostola , but present in Brachypeplus and Orthostolus . These character systems will be explored throughout the family Nitidulidae , but would not have been possible without the efforts from this research.
Pupae also yield useful species-level characters and character states, which, like larval datasets, have not been utilized in any significant phylogenetic work. The number and orientation of setiferous tubercles along the midline of abdominal tergites 2–7 are conserved in a semi-circular (5–7) and oblique manner (2–4) in B. glaber , but are found only in a transverse row (5–7) or absent (2–4) in B. ponapeus . Therefore, these characters could be coded for phylogenetic analysis separately as presence/absence data for abdominal tergites 2–4, and orientation for tergites 5–7. Likewise, the number of setiferous tubercles on each side of the midline varies between these species on tergites 5–7, with B. glaber possessing three and B. ponapeus possessing only two in each complex. The number of primary setiferous tubercles on the pronotal disc also varies between these species, with B. glaber possessing two with other secondary setiferous tubercles present, and B. ponapeus with no primary setiferous tubercles present. A potential source of species group/complex characters resides in the composition of the tubercles and processes of abdominal tergite 9. The presence of horn-like urogomphal projections in B. glaber is quite different from the complete lack of any projections in B. ponapeus . Likewise, B. glaber has complex lateral projections along the lateral margins of tergites 3–8, which is also completely lacking in B. ponapeus . These data indicate that pupal morphology may help clarify not only species-level relationships, but may also yield interesting species complexes within the genus or even characters for establishing separate monophyletic genera that were once considered Brachypeplus . At the generic level, other taxa in the Cillaeinae possess a primary seta on the developing elytra, e.g. Macrostola ; however, others lack a primary seta, e.g., Brachypeplus and Conotelus . The orientation of setiferous tubercles on the abdominal tergites also varies between genera with Conotelus and Brachypeplus sharing an oblique orientation and Macrostola with a longitudinal orientation to the midline of the body. The presence/ absence of a pleural setiferous tubercle on abdominal tergite 3 also varies among these three genera. Likewise, the shape of the eye from circular to teardrop-shaped varies as well. These and other characters would all aid in constructing a natural classification for this subfamily based on multiple datasets, including those from immatures.
Life history stage associations
The state of knowledge for immature Nitidulidae , like many Cucujoidea families, remains in its infancy with no comprehensive identification keys for larvae or pupae. Although some genera are known due to their prevalence in agricultural situations (e.g., Carpophilus , Stelidota , Epuraea , and Brassicogethes ) or occurrence in easily accessible and/or recognizable substrates (e.g., Pocadius on Lycoperdon puffball fungi, and Glischrochilus in fermenting tree sap), most genera remain unknown. In particular, generic larval descriptions of subcortical Nitidulinae and Cillaeinae are particularly depauperate, with the exception of one regional work from Japan that comprehensively covered most species and all genera from that area (Hayashi 1978). This is likely due to their cryptic and often inaccessible habitats, and inability to associate adults with corresponding larval forms. Molecular data indicated a 100% match of nucleotides for the barcode region of CO1 between adults, larvae, and pupae. All sequences have been deposited in GenBank and are available for other researchers. Thus, our rearing and molecular methods for associating immatures of B. glaber with adults of the species should provide others with a means to establish a 100% connection between life stages of this species. This same method was recently used to confirm the specific identity of larval and adult stages and host-plant associations of a Brassicogethes species in northwestern Spain (Audisio et al. 2011). Molecular methods for associating life history stages have been performed in other arthropods with similar success (Caterino & Tishechkin 2006; Slowik & Blagoev 2012); however, this is the first time these methods have been extensively used for Nitidulidae .
The description of immature forms for B. glaber provided here are the first known complete immature descriptions for this genus. Böving & Rozen (1962) included B. rubidus (an African member of the genus) in their key to mature larvae of Nitidulidae (couplet 21) and provided drawings of the mesothorax, first abdominal segment and the spiracular tube of the first abdominal segment. However, a complete description is lacking and there are no habitus or other figures provided. In addition, B. rubidus pupae were partially figured, but not fully described, in a preliminary treatment of nitidulid pupae (Rozen 1963). Thus, we feel warranted in our statement that this is the first comprehensive larval or pupal description for the genus. The pupal stage of Brachypeplus is thus far known only from the African B. rubidus (Rozen 1963) and our observations herein for B. glaber . This is significant not only for understanding the immature stages of these beetles, assessing potentially phylogenetic characters, and providing a means to identify this species, but also for developing a more robust knowledge base for understanding the biology of these beetles in confined subcortical conditions and how this biology may reflect ecological interactions with congeners (of different life stages), commensals, and food sources.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |