
Microlestes aljezurensis Ortuño & Oliveira, 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.214428 |
|
DOI |
https://doi.org/10.5281/zenodo.6173480 |
|
persistent identifier |
https://treatment.plazi.org/id/03A8F855-5F58-2955-FF0F-909DFEEEFDC7 |
|
treatment provided by |
Plazi |
|
scientific name |
Microlestes aljezurensis Ortuño & Oliveira |
| status |
sp. nov. |
Microlestes aljezurensis Ortuño & Oliveira View in CoL , new species
( Figs. 1–12 View FIGURE 1 View FIGURES 2 – 3 View FIGURES 4 – 6 View FIGURES 7 – 10 View FIGURES 11 – 12 )
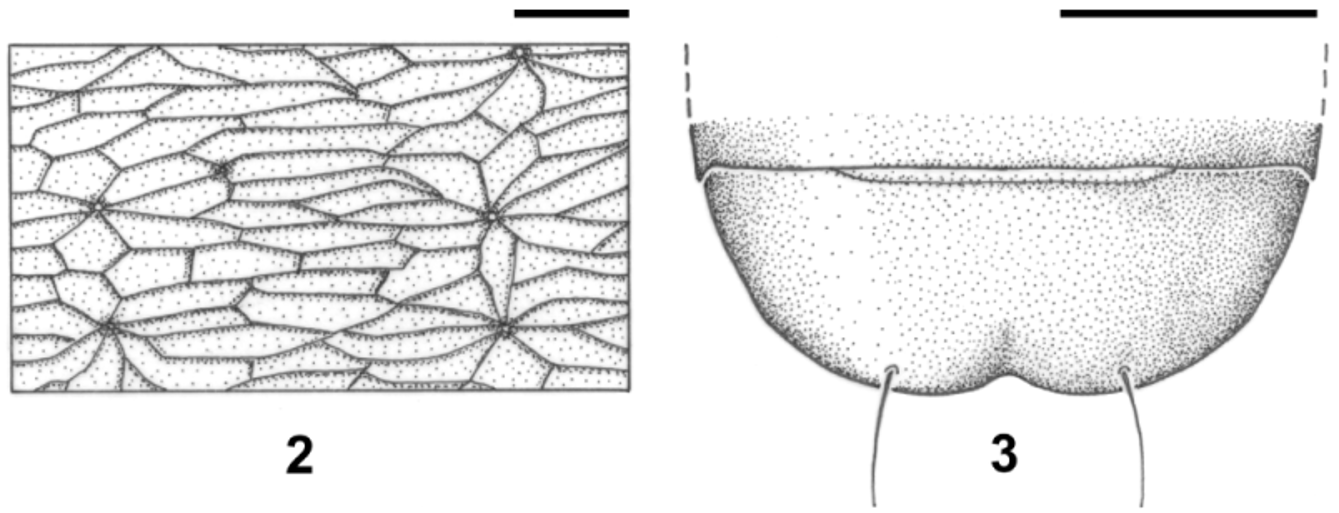
Diagnosis. Length: 2.7 to 3.3 mm. Micropterous species (wings reduced to tiny stumps). Stylized form (by keeping the proportion of the width of the elytra relative to the width of the pronotum). The elytra are short, with the apex truncated perpendicular to the elytral suture, exposing the pygidium, making it clearly visible, and partially exposing the prepygidial segment. Integument shiny black, with the appendices also dark (black femurs and tibiae, tarsi, antennae and palpi with brown-dark or almost black tones); transverse microsculpture ( Fig. 2 View FIGURES 2 – 3 ). Chaetotaxy normal for the genus.
Description. Head ( Fig. 1 View FIGURE 1 ): large, with visibly protruding eyes, whose convexity is extended into long temples that end in a thick neck. Amply extended cephalic disk, devoid of wide and deep supraocular sulci. The temples have a set of microseta. Labrum trapezoidal. The antennae, although filiform, are rather short, slightly surpassing the elytra third basal. The mandibles, as in other species of the genus, are not dorsally visible in the resting position. The maxillary and labial palpi do not offer any distinctive features.
Pronotum ( Fig. 1 View FIGURE 1 ): small, transverse (L/A ≈ 0.7), convex and cordiform, slightly wider than the head (including the eyes). The sides are curved and narrowed backwards, and sinuate near the posterior angle. Its greatest width is located at the level of the anterior marginal seta. The basal margin is sinuated at the ends and the base is almost pedunculated in the centre and visibly notched at the lateral ends, tracing an oblique line that reaches the posterior angle, which is obtuse. The anterior angles are less prominent. The basal pits are very shallow and thus poorly marked. The pronotal disk is divided by a longitudinal groove.
Elytra: elytra ( Fig. 1 View FIGURE 1 ) subconvex, broad, of suboval contour (minimum width in the humeral region, L/A ≈ 2.0); maximum width near the apex, L/A ≈ 1.15), apical truncation perpendicular to the sutural line. Obsolete striations (completely deleted or slightly shown), making the interstriae flat, and giving a smooth appearance to the elytral surface. Elytral suture separately rounded (dehiscent) at the tip. Elytral chaetotaxy: each elytron has a scutellar seta, two setigerous disk pores (no seta observed on them), one apical seta and eleven seta in the umbilical series.
Legs: prothoracic legs ( Fig. 4 View FIGURES 4 – 6 ) with no special characteristics, with two clip seta in the protibial antennal cleaning organ; males show the protarsus weakly widened. Metathoracic and mesothoracic legs with special features in males ( Figs. 5–6 View FIGURES 4 – 6 ): mesofemur with an obtuse tooth placed on the inner edge ( Fig. 5 View FIGURES 4 – 6 ); mesotibia with a distal tooth in the inner surface ( Fig. 5 View FIGURES 4 – 6 ); curved metatibia in its distal part with 4 or 5 setulose denticles giving it a clearly serrulated look in the distal half of the inner surface ( Fig. 6 View FIGURES 4 – 6 ). The females show smoother serrulated metatibias.
Abdomen: last visible abdominal male segment with a slight concavity near the distal margin, which is slightly notched ( Fig. 3 View FIGURES 2 – 3 ).
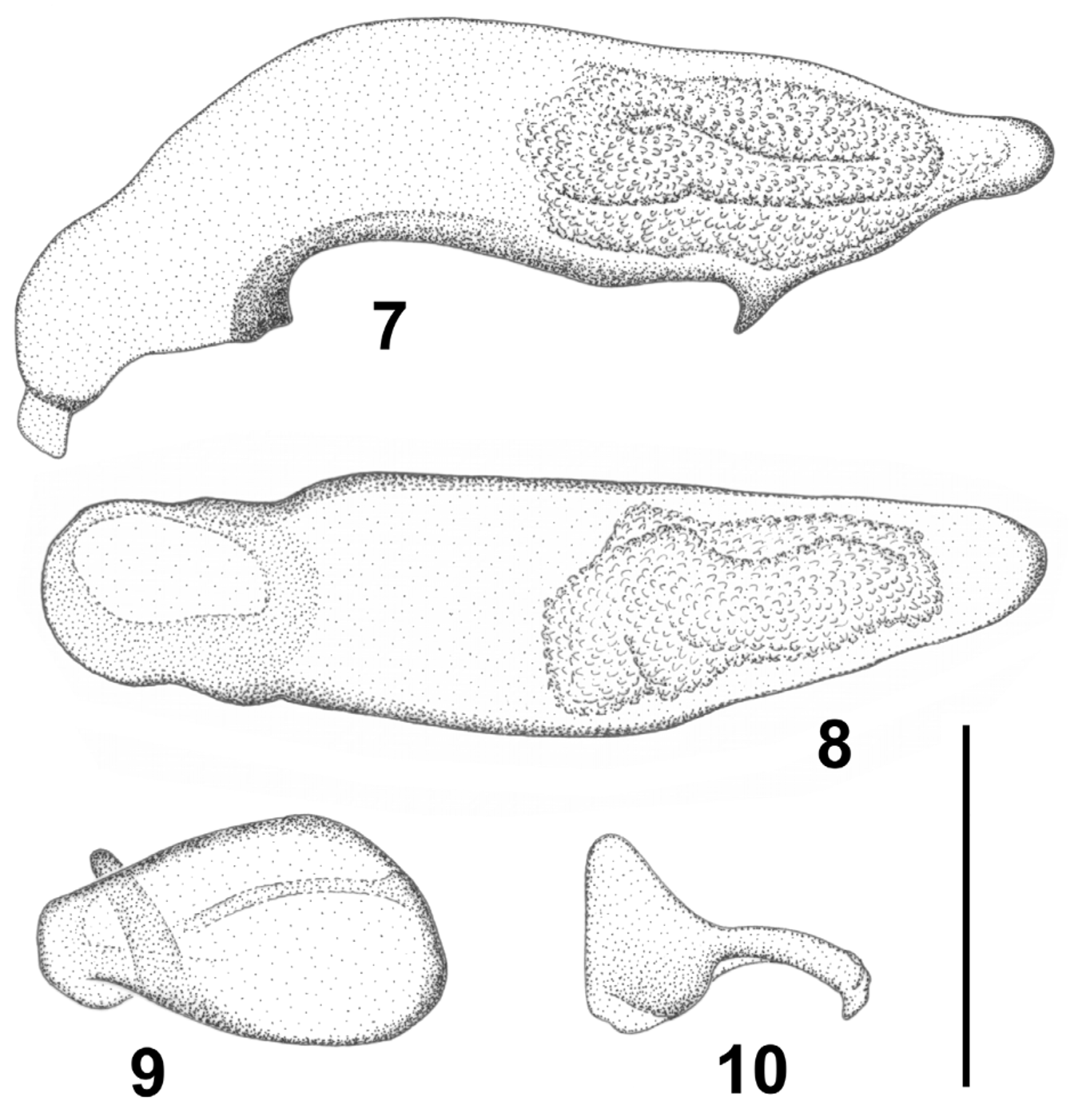
Aedeagus: median lobe ( Figs. 7–8 View FIGURES 7 – 10 ) large, very slightly convex, almost straight and with a sagittal wing; the apex is rounded, in lateral and dorsal view; there is a conspicuous retrograde denticle, harpoon like, which protrudes from the ventral surface away from the apex; internal sac without internal structures, formed solely by a bulbous structure of scaly nature. Unequal parameres: the left one of larger size and atrial aspect ( Fig. 9 View FIGURES 7 – 10 ); the right one smaller and of spatuliform aspect ( Fig. 10 View FIGURES 7 – 10 ).
Female genitalia ( Figs. 11–12 View FIGURES 11 – 12 ): external genitalia ( Fig. 11 View FIGURES 11 – 12 ) feebly sclerotized formed by dimerous IX gonopod and IX laterotergites: subtrapezoidal gonocoxite with rounded angles, especially at the distal end in whose proximity, and on the ventral surface, are inserted two thin seta; small gonosubcoxite in relation to the size of the gonocoxite, and devoid of seta; aliform laterotergite IX with several pores near the basal margin, although they do not show themselves perceptively setulated. Spermathecal complex ( Fig. 12 View FIGURES 11 – 12 ): bulky copulative bursa which leads, through a short and wide hyaline duct, to a subspherical and hyaline dilation in which ends the fine accessory gland duct; the spermatheca, thick, finger shaped, hyaline and ringed, opens next to the subspherical dilation; the base of the spermatheca has a sclerotized ring: the odd oviduct opens directly in the end of the bursa.
Type series. Holotype: 3, Poldra, Serra de Monchique, T.M. Aljezur (Algarve, Portugal), 28.V.2010, A. Oliveira leg.
Paratypes: same location and collector; 1 3, 20.IV.2007; 2 3 and 1 Ƥ, 17.V.2007, 1 3, and 1 Ƥ 25.VI.2008, 1 Ƥ 23.VII.2008, 6 3, 28 -V. 2010.
The typical series were deposited at Vicente M. Ortuño’s (VMO/AU) collection in the Departamento de Zoología y Antropología Física de la Universidad de Alcalá, and the Museo Nacional de Ciencias Naturales ( MNCN).
Etymology. The specific name “ aljezurensis ” is a tribute to the municipality of Aljezur, on whose lands this new species was found.
Biology. Microlestes aljezurensis n. sp. was found in a small area (3 squares each of 1 km x 1 km) in the foothills of Serra de Monchique ( Fig. 13 View FIGURE 13 ), on clay soils with schist slates with the following botanical characteristics: a) in oak-cork woodland, “montado” of Quercus suber L. with shrub dominated by Cistus ladanifer L.; b) in “montado” without shrub cover; c) in a strawberry tree area ( Arbutus unedo L.) accompanied by Cistus crispus L., C. ladanifer and Lavandula sp. It was also found on the margins of an irrigation pond located in a “montado” with scrubs. Usually found in open, sunny areas on paths that cross these places, and also in small gaps with grass and cracked soil.
All sites where M. aljezurensis n. sp. was located are at a low altitude (between 150 and 230 m. a.s.l.), although in two of these locations the species appeared at the highest point of the surrounding area. The captures were made during the months of April, May and June, by installing pitfall traps. While performing capture transects, the specimens were localized to the following temperatures: air temperature (max: 23.4 ºC–min: 12.6 ºC), temperature of the soil surface (max: 33.9 ºC–min: 13.4 ºC) and soil temperature at 15 cm depth (max: 28.8 ºC–min: 17.5 ºC). The grid cells surrounding the three cells where M. aljezurensis n. sp. was encountered were prospected on several occasions, but in all cases the results were negative.
Other species of Carabidae that coexist in the same UTM grid cells where M. aljezurensis n. sp. is found are indicated in Table 1.
SPECIES Nº of specimens Carabus (Macrothorax) rugosus celtibericus Germar, 1824 10 Carabus (Mesocarabus) lusitanicus lusitanicus Fabricius, 1801 3 Carabus (Rhabdotocarabus) melancholicus costatus Fabricius, 1798 1 Notiophilus quadripunctatus Dejean, 1826 1 Trechus (Trechus) fulvus Dejean, 1831 5 Trechus (Trechus) obtusus Erichson, 1837 10 Elaphropus (Tachyura) parvulus (Dejean, 1831) 3 Tachys (Paratachys) bistriatus (Duftschmid, 1812) 10 Bembidion (Phyla) tethys Netolitzky, 1926 3 Abacetus (Astigis) salzmanni (Germar, 1824) 7 Pterostichus (Oreophilus) paulinoi vanvolxemi Putzeys, 1874 1 Steropus (Sterocorax) globosus ebenus (Quensel, 1806) 90 Olisthopus hispanicus Dejean, 1828 1 Paranchus albipes (Fabricius, 1792) 3 Platyderus sp. 5 Amblystomus escorialensis Gautier des Cottes, 1866 1 Amblystomus niger (Heer, 1841) 22 Carterus (Carterus) fulvipes (Latreille, 1817) 1 Dixus sphaerocephalus (Olivier, 1795) 1 Harpalus (Harpalus) attenuatus Stephens, 1828 1 Stenolophus teutonus (Schrank, 1781) 3 Chlaenius (Chlaenius) festivus velutinus (Duftschmid, 1812) 1 Masoreus wetterhallii (Gyllenhal,1813) 1 Mesolestes (Mesolestes) scapularis (Dejean, 1829) 2 Microlestes abeillei abeilleiI (Brisout de Barneville, 1885) 2 Microlestes corticalis (Dufour,1820) 2 Microlestes luctuosus luctuosus ( Holdhaus, 1904) 9 Microlestes negrita negrita Wollaston, 1854 8 Syntomus obscuroguttatus (Duftschmid, 1812) 6
Taxonomy. The discovery of M. aljezurensis n. sp. increases the number of Microlestes species found in the Ibero-Balearic area to 16. Identification of most of these species is extremely difficult due to their great resemblance in external morphology. Males of different species may sometimes be distinguished by the curvature of the mesotibia and metatibia and the presence of spinous processes in distal position, the different degree of dilation of the protarsus, or the presence of indentations and impressions more or less ornamented in the last abdominal sternite. However, the aedeagus is the best structure for identifying these species, whereas the female genitalia have a sclerotized ring shaped structure (= annulus receptaculi; Holdhaus 1912) that may differ among species ( Holdhaus 1912; Mateu 1953, 1960, 1971a, 1974, 1976; Coulon 1998). However, our opinion is that the use of the female genitalia is not always sufficient for identifying all of the species found in the Ibero-Balearic area. An example is the extraordinary resemblance between the female genitalia of M. gallicus Holdhaus, 1912 , M. seladon Holdhaus, 1912 and M. negrita (Wollaston, 1854) , or between M. abeillei (Brisout de Barneville, 1885) and M. fissuralis (Reitter, 1901) (see Coulon 1998). For this reason, we decided to develop a key to the Iberian taxa without using the characteristics of the female genitalia.
To date, the only keys for identifying Iberian-Balearic Microlestes are those developed by De la Fuente (1927), which are now outdated. Therefore, addressing the identification of these species could only be done by adding to the previous book the ancient monograph of Holdhaus (1912) or works of a regional nature, such as the Faune de France ( Jeannel 1942; Coulon 1998) or that of Morocco ( Antoine 1962). For this reason, we thought it appropriate to prepare a key containing all of the Ibero-Balearic species to facilitate future study of Microlestes in this geographical area.




| MNCN |
Museo Nacional de Ciencias Naturales |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |