
Austrodontella monticola, Babenko, Anatoly & Minor, Maria, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3974.1.9 |
|
publication LSID |
lsid:zoobank.org:pub:5E90A96F-7450-4699-ACB7-E7E146F589F5 |
|
DOI |
https://doi.org/10.5281/zenodo.5680542 |
|
persistent identifier |
https://treatment.plazi.org/id/03A987C9-B258-720F-56FF-FBDA17647D92 |
|
treatment provided by |
Plazi |
|
scientific name |
Austrodontella monticola |
| status |
sp. nov. |
Austrodontella monticola sp. nov.
Figs 1–10 View FIGURES 1 – 10
Type locality. New Zealand, South Island, Central Otago, Pisa Range, herbaceous snowbank, in the soil, 44°52'25.9536"S, 169°10'31.9476"E, 1808 m a.s.l.
Type material. Holotype, male, locality as above, 18 February 2014, M. Minor leg.
Paratypes: 4 females and 5 males, same sample as holotype; 1 female, Pisa Range, alpine cushionfield, bare ground with some lichen near Raoulia sp. cushion, in the soil, 44°52'11.1"S, 169°10'9.195"E, 1797 m a.s.l., 18 February 2014; 1 male, Pisa Range, alpine bog, in moss, 44°51' 59.004"S, 169°9'30.6324"E, 1688 m a.s.l., 18 February 2014; 1 juv., Central Otago, Old Man’s Range, alpine cushionfield, bare ground with lichen near Raoulia sp. cushion, in the soil, 45° 19' 7.6038"S, 169° 12' 11.4876"E, 1655 m a.s.l., 17 February 2014, all M. Minor leg.
Diagnosis. Odontellidae with three anal spines set in a line. PA O amoeboid, usually with four lobes. 5+5 subequal ocelli present. Ant.4 with domed apical swelling but without eversible bulb; subapical organite, microsensillum and thick curved sensilla present. Buccal cone short. Labial palp with 5 minute spinules each set on low papillae and 2 proximal short setae. Labrum with 2/44 setae. Maxillary palp present, simple (i.e. with single hair). Dorsal terga, excepting Th.2–3 [with m 4 and m 6 (= s) present], devoid of setae m. Furca complete, dens with (3)4 setae, mucro short with tooth on outer lamella. Ti.1–3 without setae M and A 4 (17–17–16(15) setae as a whole), seta A 1 long but not clavate.
Description. Size 0.9–1.2 mm. Body rather narrow and long, general habitus is similar to Frieseinae rather than Odontellidae . Granulation coarse and uniform with 4–8 granules between setae p 1 on Abd.5. Colour spotted pale bluish (alcohol), ocular field darker.
Antennae almost as long as head length. Ant.1–3 with 7, 11, and 16 ordinary setae, respectively. AO on Ant.3 with 5 sensilla as usual for most Poduromorpha: two small rounded internal sensory rods (s 2 and s 3), two longer external sensilla (s 1 and s 4), and lateral microsensillum (s 5) ( Fig. 3 View FIGURES 1 – 10 ). Ant.4 with a clear domed apical swelling, eversible vesicle absent; subapical dorsal organite, microsensillum and some thick curved subequal sensilla present; the number and position of the latter slightly variable: three dorsal (probably, s 1, s 4 and s 2 (or s 5)) and usually 4 lateral (s 7, s 8, s 9, and X) ( Fig. 3 View FIGURES 1 – 10 ), few specimens with additional fifth sensillum in lateral group have been seen. Ventral file on Ant.4 with about 10 straight, short and slightly clavate setae and two blunt sensilla. 5+5 ocelli present, rather large and subequal. PA O as long as or slightly longer than nearest ocellus, amoeboid in shape and with four or more unequal lobes ( Figs 4–5 View FIGURES 1 – 10 ), in few cases the fourth lobe short ( Figs 6-7 View FIGURES 1 – 10 ) but always visible in all available specimens. Buccal cone rather short. Mouthparts poorly sclerotized and similar to those in A. cassagnaui (cf. fig. 11B in Deharveng 1981): mandibles strongly reduced, styliform, maxillary head with a number of small teeth distally, cardo fused with fulcrum, EPP and IPP of fulcrum of similar length ( Fig. 8 View FIGURES 1 – 10 ). Maxillary palp present, simple and without sublobal setae. Labrum with 2/44 setae, medial pair of distal row modified, curved and pressed down to labral edge. Clypeal field with 3 setae: one above two prelabral setae and two near lateral sides of labrum (thus, it’s possible that 4 prelabral setae exist in fact). Labial palp with 5 minute spinules, each set on low papillae, and 2 proximal setae ( Fig. 2 View FIGURES 1 – 10 ). Basomedial (submentum) and basolateral (mentum) parts of labium with 4 and 6 setae, respectively; 3+3 postlabial setae present along linea ventralis, medial one being the longest.
Dorsal chaetotaxy as on Fig. 1 View FIGURES 1 – 10 , variations not frequent. Ordinary setae rather long, especially on abdominal tip, usually smooth and pointed, rarely slightly serrated on last abdominal terga. Sensorial setae (s) poorly differentiated, slightly thinner than and about as long as adjoining ordinary setae, their number and position as usual: 22/11111, lateral ms present on both Th.2 and Th.3. Main characteristics of chaetotaxy: head with seta a 0 present and only two setae in c -row; Th.2–3 with only two setae (m 4 and m 6 (s)) in m -row; only two axial pairs of setae on all terga from Th.2 to Abd.5. Ventral chaetotaxy of abdominal segments as on Fig. 9 View FIGURES 1 – 10 . Thoracic sterna devoid of setae.
Number of setae on legs as in Table 1 View TABLE 1 . Ti.1–3 without setae M and A 4 on all legs, Ti.3 also lacking seta B 6 (and sometimes B 7), A 1 rather long (about as long as inner edge of unguis) and pointed. Unguis without or with hardly visible tooth on basal third of inner edge, lateral ones invisible. Unguiculus absent. Ventral tube with 3+3 lateral setae, two of which set more distally, one additional seta always present on sternum of Abd.1 at each side of the tube base. Tenaculum with 3+3 teeth, sometimes the fourth additional tooth present asymmetrically. Furca complete with all parts present ( Fig. 10 View FIGURES 1 – 10 ). Manubrium as a rule with 9+9 posterior setae and with 4+4 setae on laterobasal lobes. Dens with (3)4 posterior setae of similar length, mucro about 1/3 of dens (mucro: dens = 1: 2.5– 3.4), with triangular tooth on outer lamella. Each anal valve with three setae hr, mature males with 2 spine-like setae on each of paired valve ( Fig. 9 View FIGURES 1 – 10 ), which are probably the same as deux elements piliformes plats et larges in anal region of A. trispina mentioned by Massoud (1967). Anal spines on Abd.6 strong, set in a line on high granulated papillae, middle one longer.
Variations. Despite the relative shortage of the material, two notable types of variation were detected within it. The bulk of the type material which originated from a single sample [Pisa Range, snowbank, 10 specimens in total] lacks two B -setae on Ti.3, whereas three other specimens from other soil cores possess 6 setae in the B -whorl of Ti.3, the latter is more common condition for Poduromorpha. The number of tibiotarsal setae for any species is normally considered stable.
Apart from this, one female among the types was characterised by a clearly reduced furca with shortened dens bearing only two setae and small mucro almost lacking tooth on outer lamella. This condition appears to correlate with less sclerotized mouthparts, more flattened buccal cone, and shortened sensilla on Ant.4, as well as almost all setae shortened, including tibiotarsal ones. Similar although less pronounced modifications were found in the single juvenile available (Old Man’s Range, cushionfield). Most probably it is the result of ecomorphosis as understood by Cassagnau (1956) —temporary changes in some morphological traits in unfavourable environmental conditions.
Etymology. The specific epithet refers to the high alpine zone where the species was found.
Affinities. The new species is similar to A. trispina . Based on the existing descriptions ( Womersley 1935; Massoud 1967) of the latter, these two species can be distinguished by their sizes (0.9–1.2 mm in A. monticola sp. nov. vs 0.55 mm in A. trispina ), number of lobes in PA O (three in A. trispina vs more lobes in A. monticola sp. nov.), tegument granulation, which is said to be fine in A. trispina (le grain tégumentaire est très fin according to Massoud (1967, p. 81)), and mucro length (equal to dens in A. trispina vs ~1/3 of dens in A. monticola sp. nov.). As all these traits appear, however, to be rather variable, we have studied two of Womersley’s syntypes of A. trispina (slides I22382 & I22383) obtained from the South Australian Museum in Adelaide. The condition of both syntypes is good and our finding may be summarised as following: granulation of tegument of the types was not as fine as stated by Massoud (1967) with about 5–6 granules between setae p 1 on Abd.5 ( Fig. 11 View FIGURES 11 – 12 ) which is within a range typical of A. monticola sp. nov.; mucro in both specimens appeared to be not equal to dens but only 0.5–0.7 of dens’ length which further removed a difference between the two species. Nevertheless, A. monticola sp. nov. and A. trispina clearly differ in tibiotarsal chaetotaxy, which is strongly reduced in the latter species with only 10–10–9 setae on Ti 1–3, respectively ( Figs 12–13 View FIGURES 11 – 12 View FIGURES 13 – 14 ). The difference in the relative lengths of the middle unpaired and the lateral paired anal spines in A. trispina ( Fig. 14 View FIGURES 13 – 14 ) compared to the new species where these three spines are similar in length is another distinguishing character.
The second described species, A. cassagnaui , with a fuller original description, can be separated because it also has fewer tibiotarsal setae (15–15–14 setae in A. cassagnaui , including two distinctly clavate ones, vs 17–17– 16(15), all pointed in the new species). Other constant differences can be listed: 6 vs 8 labral setae, dens: mucro ratios as 2.1 vs 2.5–3.4 in A. monticola sp. nov., seta a 0 on head (absent in A. cassagnaui vs present in A. monticola sp. nov.), setae p 2 on Abd.1–3 (sometimes absent in A. cassagnaui vs always present in A. monticola sp. nov.).

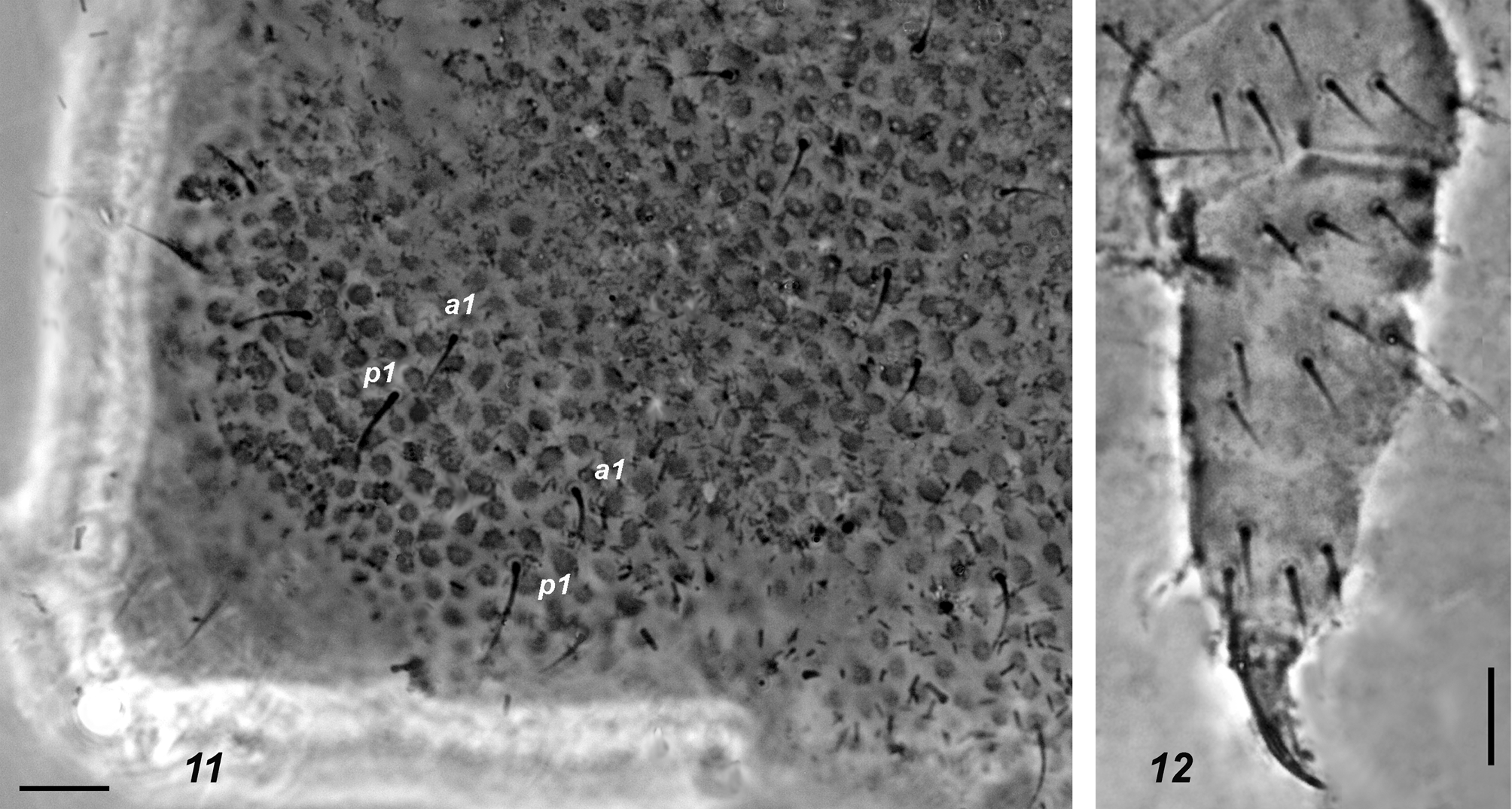
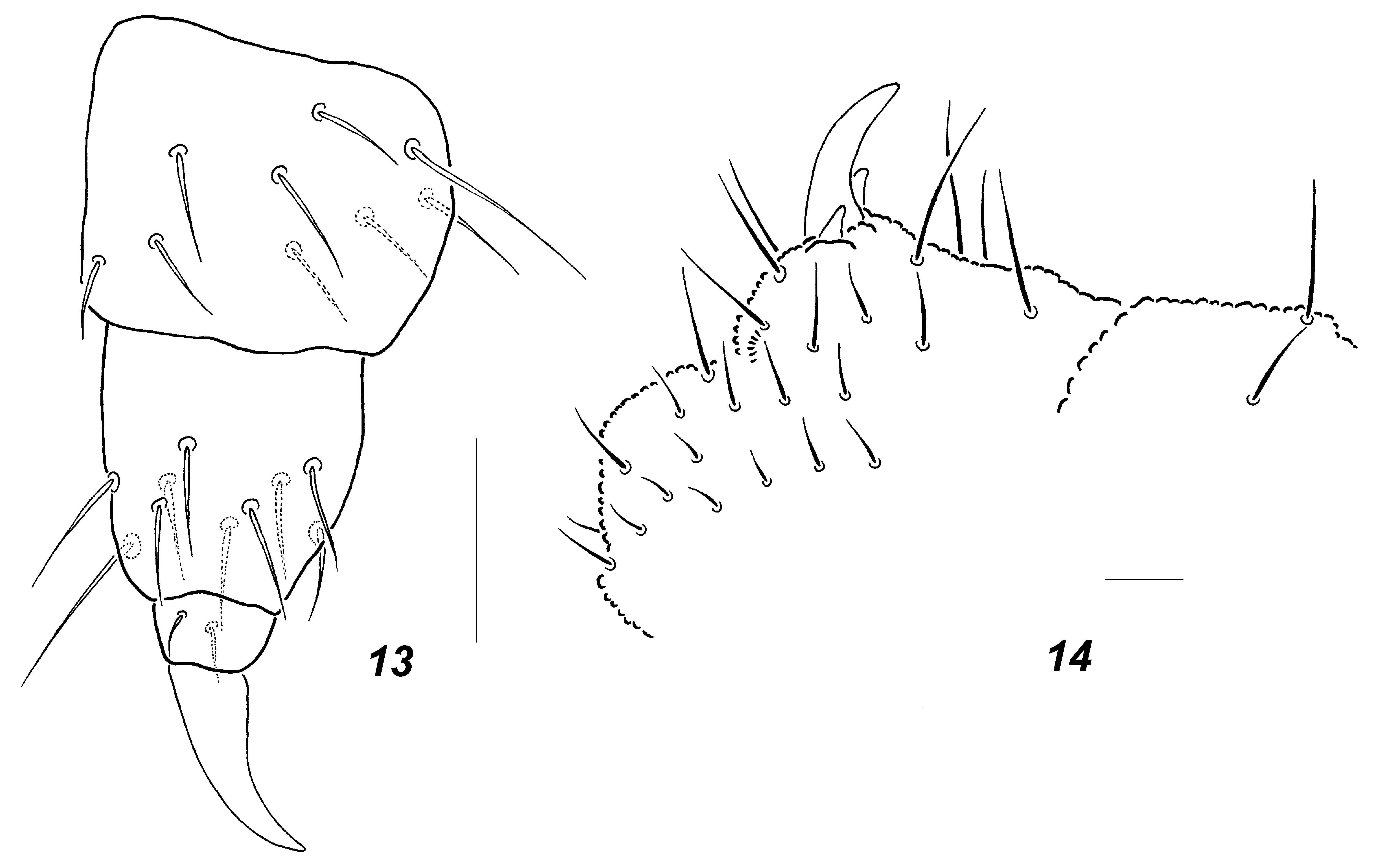
TABLE 1. Leg chaetotaxy of A. monticola sp. nov.
| Upper subcoxa | Subcoxa | Coxa | Trochanter Femur | Tibiotarsus |
|---|---|---|---|---|
| Leg 1 1 | 0 | 3 | 5 11–12 | 17 |
| Leg 2 2 | 2–3 | 7–8 | 5 11 | 17 |
| Leg 3 2 | 2–4 | 7–8 | 4–5 9–10 | 15–16 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |