
Dagonodum mojnum, Benjamin Ramassamy, 2016
|
publication ID |
https://doi.org/10.1111/zoj.12418 |
|
DOI |
https://doi.org/10.5281/zenodo.4426021 |
|
persistent identifier |
https://treatment.plazi.org/id/03AB87AC-4F28-FFA6-D51F-FE9BFA3EF97F |
|
treatment provided by |
Plazi |
|
scientific name |
Dagonodum mojnum |
| status |
sp. nov. |
DAGONODUM MOJNUM SP. NOV.
Holotype. MSM1001x, a partial skeleton including a fragmentary cranium, the left periotic and tympanic, both mandibles, 75 isolated teeth, five cervical vertebrae including the atlas and the axis, two thoracic vertebrae, several fragmentary ribs, the basihyal and left thyrohyal.
Diagnosis. Dagonodum mojnum is a medium-sized ziphiid with an elongated rostrum (approximately 70% of the total condylobasal length). It differs from all odontocetes except ziphiids in having the vertex bearing a strong premaxillary crest; the enlargement of the apical mandibular tooth, and the reduction of the dorsal keel on the posterior process of the periotic.
It shares with Anoplonossa forcipata and Berardius the presence of a pair of apical and subapical tusks; with the genera Messapicetus and Ziphirostrum the medial fusion of the premaxillae dorsal to the mesorostral groove on the rostrum anterior to the premaxillary sac fossae; and with Aporotus , Beneziphius , Messapicetus and Ziphirostrum the presence of a prenarial basin laterally margined by the maxilla.
Dagonodum mojnum differs from all other Ziphiidae based on the following unique combination of characters: nasal anteroposteriorly elongated and trapezoid in dorsal view; left nasal projecting into the frontal posteromedially; presence of a triangular depression on the anteroventral surface of the pars cochlearis; Eustachian outlet approximately level with the dorsalmost margin of the posterior portion of the involucrum in medial view; presence of an incipient anterior spine on the tympanic bulla; and presence of two anterodorsally directed pairs of mandibular tusks.
It further differs from all Berardiinae in having a moderate transverse constriction in the ascending process of the premaxilla, a larger premaxillary crest equivalent in transverse width to the nasal and in lacking a posterior rounded protuberance formed by the frontals and/or interparietal on the vertex; from the Hyperoodontinae in the lack of a deep anteromedial excavation of the nasals; and from the Ziphiinae in the lack of a premaxillary sac fossa overhanging the maxilla laterally.
Dagonodum mojnum differs from Globicetus hiberus , Imocetus piscatus and the genus Tusciziphius in lacking an extreme ossification and fusion of the bones on the trapezoidal vertex and the absence of a dorsal prominence on the premaxillae along the rostrum (not present in I. piscatus ); from Nenga meganasalis and Xhosacetus hendeyesi in having a narrower nasal not occupying most of the vertex; from Pterocetus benguelae in having a shallower antorbital notch with a less developed preorbital process; from Tasmacetus shepherdi in having the premaxillary crest anterolaterally orientated and the external surface of the nasal facing dorsally, not progressively sloping ventrally; from Aporotus dicyrtus , A. recuvirostris, Beneziphius brevirostris and Ziphirostrum marginatum in lacking a strong thickening of the maxilla at the base of the rostrum and in having a prominential notch (condition unknown in Beneziphius ); from Aporotus in having the premaxillae dorsally roofing the mesorostral groove fused; and from Beneziphius in lacking the extension of the medial fusion of the premaxillae more posteriorly until the premaxillary sac fossae.
Dagonodum mojnum differs from Messapicetus in having a less pronounced transverse constriction in the ascending process of the premaxilla characterized by U-shaped margins (ratio between transverse width of the premaxillary crest and minimum transverse width of the ascending process equals 0.72); an anteroposteriorly longer nasal as wide as long (ratio between median length and total width equals 0.53); and a posteromedial projection of the nasal into the frontal and in lacking transversely compressed alveoli.
Etymology of species name. From the word ‘mojn’ meaning ‘morning’ in South Jutlandic. Originally of German origin, this is still used as a greeting for ‘hello’ and ‘goodbye’ in Southern Jutland.
Horizon and locality. The locality is situated 1.5 km north of the town of Gram, Southern Jutland, Denmark (55 ° 18 l 25.67 ″ N, 9 ° 3 l 32.51 ″ E; Fig. 1 View Figure 1 ). The mollusc fauna found in association with MSM1001x was identified. The high percentage of Carinastarte vetula reimersi (34% of the mollusc species identified) and the co-occurrence of the species Turitella cochlias and Ringicula buccinea indicates that the specimen MSM1001x was situated in Assemblage Zone III or IV described by Rasmussen (1966). However, the species T. tricarinata was also identified, whereas it was not reported by Rasmussen (1966) in Zones III and IV of the Gram claypit. The co-occurrence of these three mollusc species is reported in other localities of the Gram Formation, in the Odderup borehole belonging to Zones III and IV, and in the borehole at Brande Brickworks belonging to assemblage Zone V ( Rasmussen, 1966). The specimen is therefore identified as belonging to the Zones III, IV or V.
From a magnetostratigraphic viewpoint, the occurrence of a reverse polarity zone suggests a minimum age of 9.9 Mya for the Assemblage Zones of the Gram Formation and the consistent occurrence of the dinoflagellate cysts Hystrichosphaeris obscura, Spiniferites solidago and Labyrinthodinium truncatum indicates a late Tortonian age for the upper part of the Gram Formation ( Beyer, 2005; Piasecki, 2005). MSM1001x can therefore be dated to the mid- to late Tortonian, c. 9.9–7.2 Mya.
Palaeoenvironment and depositional setting. The Gram Formation was deposited in a marine environment at a depth between 50 and 100 m, as indicated by the marine flora associated with terrestrial material, foraminifera and the marine fauna ( Rasmussen, 1958, 1966; Laursen & Kristofferson, 1999; Dybkjaer & Piasecki, 2010). Fine-grained storm beds similar to the extant coastal layers in the North Sea suggest a progradation of the shoreline ( Rasmussen, 2005).
The fauna is typical of a coastal environment with 128 mollusc species and six decapod species ( Fraaije, Hansen & Hansen, 2005; Schnetler, 2005). They are associated with shark remains, among others the predator Carcharocles megalodon ( Bendix-Almgreen, 1983) . Cetaceans are also represented by at least two baleen whale species, Uranocetus gramensis ( Steeman, 2009) and Tranatocetus argillarius ( Roth, 1978; Gol’din & Steeman, 2015) and a delphinoid species ( Hoch, 2004).
Remarks on taphonomy and ontogeny. The head of MSM1001x was found with a north-east–south-west orientation. The cranium was lying with the ventral side up and was covering the mandibles both in the same orientation. Teeth were found out of their sockets, but close to the mandibles. Postcranial elements were disarticulated under and around the head of MSM1001x. Their position does not provide information about the original articulation between elements.
The individual is mature as indicated by the strong fusion of the vertebral epiphyses on the cervical and thoracic vertebrae and the extensive wear observed on the apical tusk. The dorsomedial fusion of the premaxillae on the rostrum along the mesorostral groove is consistent with this interpretation, given that an immature individual of the species Messapicetus gregarius lacks this feature ( Bianucci et al., 2010).
Cranium and mandibles
Overview. The general dimensions of the skull of MSM1001x were estimated based on comparisons with the other long-snouted beaked whales Ninoziphius platyrostris , Messapicetus gregarius and M. longirostris . The condylobasal length is estimated to be 970 mm (minimum) and the preorbital skull breadth at least 311 mm. The mandible length is estimated to 843 mm. Based on those estimates, D. mojnum is a medium-sized ziphiid smaller than Messapicetus spp., but similar in proportions to N. platyrostris . The skull and mandibles of D. mojnum were reconstructed by comparison with those fossil ziphiids ( Fig. 2 View Figure 2 ). The preserved portion of the cranium comprises the skull roof, facial area and the detached rostrum, the rostrum missing c. 100 mm at its proximal portion ( Figs 3–7 View Figure 3 View Figure 4 View Figure 5 View Figure 6 View Figure 7 ). The rostrum is relatively well preserved thanks to its high bone density, but a dorsoventral compression caused the vertex to fracture and collapse anterolaterally toward the left ( Figs 3 View Figure 3 , 5 View Figure 5 ), shifting the maxilla and frontal forward. The earbones are rather complete, but were found detached from the rest of the cranium ( Figs 8 View Figure 8 , 9 View Figure 9 ). The mandibles are missing most of the ramus ( Fig. 10 View Figure 10 ). Skull measurements are provided in Table 1 View Table 1 . Unless otherwise mentioned, the description refers to the left part of the skull.
Premaxilla. Along the rostrum, the premaxilla is more elevated than the maxilla in lateral view. The suture between the two bones is not marked. Posterior to the rostral apex, the premaxillae are dorsomedially fused, with a thin groove marking the point of contact ( Fig. 3 View Figure 3 B–D). The proximal rostral region where the fusion first occurs is not preserved. On the rostrum base, the dorsalmost surface of the premaxilla is more ventral than the maxilla. The premaxilla, maxilla and vomer together form the prenarial basin ( Fig. 6 View Figure 6 ).
Ventromedially along the rostrum, a palatal trough is developed between the two alveolar grooves. The trough excavates both the maxilla and the premaxilla, and is dorsally roofed by the vomer. The suture between the premaxilla and maxilla in this region is often hard to follow. At the rostral apex, the premaxilla alone forms the palatal trough. On the cranium, the left premaxilla is artificially shifted anterolaterally on the left. The oval premaxillary sac fossae are weakly depressed, not as excavated as in Ziphius cavirostris. It is also asymmetrical with the left fossa being 77% the transverse width of the right one. The contact between the premaxillary sac fossa and the maxilla is visible only on the right side. The premaxillary sac fossa is as elevated dorsally as the maxilla, but this might be exaggerated by the deformation. The dorsal surface of the ascending process of the premaxilla is longitudinally concave. It is unclear if a posterodorsal vertical portion was present as in Ziphirostrum , or if it smoothly reached the vertex as in Berardius . The ascending process is transversely constricted; in dorsal view, the lateral margin of the ascending process is concave. The ratio between the transverse width of the left premaxillary crest (45 mm) and the minimum transverse width of the ascending process (32 mm) is 0.72. Due to the deformation, the thick and rounded premaxillary crest is detached from the frontal and the nasal. However, the orientation of the suture between the left frontal and the premaxillary crest is still directed anterolaterally ( Figs 5 View Figure 5 A–7).
Posteriorly to the crest, a narrow portion of the maxilla follows the original margin of the premaxilla and thus confirms the direction.
Maxilla. The following description is based on the right maxilla. Along the rostrum, the alveolar groove is well defined except on the anteriormost part of the rostrum ( Fig. 11 View Figure 11 C). There are 24 alveoli on each side, but the posteriormost portion of the tooth row is missing. At the base of the rostrum, there is a maxillary tubercle. Compression-related fracturing caused part of this structure to be displaced backwards, thus leading to artificial closure of the antorbital notch. The maxillary tubercle is limited anteromedially by a deep prominential notch. Medial to the antorbital notch, no rostral maxillary crest is detected. Medial and anteromedial to the prominential notch, the maxilla delimits the dorsoventrally thin lateral edge of the prenarial basin ( Figs 5 View Figure 5 , 6 View Figure 6 ).
Palatine. The palatine is not preserved, but its area of contact with the maxilla, marked by grooves along the bone surface, is still visible and extends anterior to the level of the antorbital notch ( Fig. 4 View Figure 4 A–C). The position of the palatine relative to the pterygoid sinus fossa remains unknown.
Nasal. The two nasals were originally joined and hexagonal, occupying more than half the dorsal surface of the vertex. The left nasal is dorsally well preserved and robust, anteroposteriorly elongated (49 mm long and 36 mm wide mediolaterally) and pointed anteriorly ( Figs 5 View Figure 5 , 7 View Figure 7 ). The premaxillary crest and the nasal are no longer in contact because of the deformation, but the original contact between the two bones can still be assessed based on the suture visible on the lateral surface of the nasal. It was short, extending only along 39% of the anteroposterior length of the nasal in dorsal view.
Dorsally, the posteriormost part of the left nasal is projected posteriorly into the frontal. The original outline of the posterior margin of the right nasal is harder to follow, but was probably also projected posteriorly. A deep fracture separates the dorsal and posteroventral parts of the nasals, but this did not affect the original contact of the nasals with the frontals. The nasals are also joined and posteromedially projected on their posteroventral part ( Fig. 7 View Figure 7 B, C).
Ethmoid. The mesethmoid part of the ethmoid bone forms a straight ridge anterior to the level of the right antorbital notch, at the base of the mesorostral groove ( Fig. 6 View Figure 6 ). The vertical plate of the mesethmoid anteroventrally margins the nasal.
Vomer. The vomer forms the floor of the mesorostral groove and of the prenarial basin. The vomer is not thickened in the mesorostral groove. It is also visible ventromedially, forming the medial floor of the palatal trough.
Jugal. A small part of the right jugal is visible ventrally, underlying the frontal at the level of the antorbital notch ( Fig. 4 View Figure 4 A–C).
Frontal. Dorsally on the right side of the cranium, the frontal participates in the formation of the dorsal roof of the orbit and the anterior part of the temporal fossa ( Figs 3 View Figure 3 A–C, 5A–C). On the vertex in dorsal view, the frontal is shattered, but its suture contact with the maxilla and the nasal is visible. The dorsal exposure of the frontal on the vertex is rectangular, transversely wider than anteroposteriorly long and smaller than the nasal exposure. The surface of the frontal does not form a rounded striated protuberance as observed in Antorbital notch
Berardius ( Moore, 1968) . The nasofrontal suture is anteriorly concave due to the posteromedial projection of the joined nasals ( Fig. 7 View Figure 7 ).
Interparietal. The interparietal could not be identified. If present, it is squashed in the region of the vertex. Even in extant cetaceans, this bone is hard to distinguish from the supraoccipital ( Mead & Fordyce, 2009).
Parietal. The parietal bone is shattered and its outline hard to distinguish. The posterodorsal portion of the temporal fossa is rounded, not marked by a notch posteriorly as observed in Hyperoodon ampullatus (e.g. specimens SNM CN1; MSNUP 268; MNHN A3236).
Supraoccipital. The dorsal part of the supraoccipital is preserved. Along the medial surface of the supraoccipital, a strong external occipital crest is visible posteriorly, closer to the original position of the foramen magnum, the latter not being preserved on the specimen. Each lobe of the braincase is hemispherical, slightly elongated anteroposteriorly and separated by the cruciate eminence.
Periotic. The left periotic is preserved ( Fig. 8 View Figure 8 ). Measurements of the periotic are provided in Table 2 View Table 2 . The anterior process of the periotic is slender for a ziphiid without a marked mediolateral thickening. In ventral view, it bears a rounded anterior margin and an elliptical anterior bullar facet. In ventral view, the anterior process is bounded posteriorly by a sulcus separating the lateral tuberosity and the anterior process ( Fig. 8 View Figure 8 D, H). This separation corresponds to the anteroexternal sulcus described by Fordyce (1983), but on the holotype of D. mojnum it develops more ventrally and remains visible in dorsal view. No parabullary sulcus is detected. Dorsally, the anterior process bears one exostose at the anteroventral base of the cochlea. The lateral tuberosity is rounded, transversely shorter than in extant ziphiids. It bears a wide mallear fossa that is mediolaterally elongated. A deep fossa for the stapedius muscle is located posteriorly to the fenestra ovalis laterally to the fenestra rotunda. The fenestra ovalis is elliptical, slightly compressed lateromedially. The fossa for the stapedius muscle extends posterodorsally to the facial sulcus. The facial sulcus is situated laterally to the fossa for the stapedius muscle. The facial sulcus is anteriorly connected to the distal opening of the facial canal. The distal opening of the facial canal is smaller than the fenestra ovalis, positioned slightly more posteriorly.
The anterior base of the posterior process widens transversely from anterior to posterior. It is more squared than rounded. In lateral view, the dorsal margin of the posterior process is regularly convex. In ventral view, the posterior process bears a longer posterolateral tip and its posteromedial margin is convex. Ventrally, the posterior bullar facet is not abruptly anteroposteriorly widened and is therefore not as much fan-shaped as in extant ziphiids.
The pars cochlearis is elevated medially, anteriorly shifted and square-shaped due to its straight anteromedial corner ( Fig. 8 View Figure 8 A). The wide internal acoustic meatus is separated from the aperture for the endolymphatic duct by a crest that is medially elevated, but not developed into a cochlear spine. The internal acoustic meatus is dorsoventrally elongated. Inside the internal acoustic meatus, the dorsal vestibular area is separated from its ventral counterpart by a transverse crest developed anteroposteriorly. The ventral vestibular area contains the proximal opening of the facial canal and the foramen singulare. The dorsal vestibular area contains the spiral cribriform tract and the area cribrosa media. The proximal opening of the facial canal is the most anterior opening of the internal acoustic meatus. It lays posterodorsally to the foramen singulare. The spiral cribriform tract is rounded, bigger than the area cribrosa media. The spiral cribriform tract lies anteriorly to the area cribrosa media.
The aperture for the vestibular aqueduct is located posteromedially to the internal acoustic meatus. The vestibular aqueduct is squared and larger than the aperture for the cochlear duct. The aperture for the cochlear aqueduct is located more ventrally than the internal acoustic meatus and is closer to the vestibular aqueduct than to the fenestra rotunda.
The fenestra rotunda is visible on the posteroventral surface of the pars cochlearis. It has a transversely elongated crescent shape with a thickened posteromedial border. A triangular depression is present on the anteroventral surface of the pars cochlearis ( Fig. 8 View Figure 8 D, H). The posterior point of the depression is orientated posteromedially toward the fenestra rotunda. A depression in the same area is present in Mesoplodon mirus (USNM 504612, USNM 550351, USNM 572961) and M. bidens (MNHN 1975.112, SNM CN5x), but is more anteroposteriorly elongated, not wide and triangular as in the holotype of D. mojnum .
Tympanic bulla. The dorsal part of the outer lip of the left tympanic bulla is missing, together with the sigmoid process and the posterior process ( Fig. 9 View Figure 9 ). Measurements are provided in Table 2 View Table 2 . The bulla is heart-shaped in ventral view with two asymmetric posterior prominences: the inner posterior prominence is transversely compressed and narrower than the outer posterior prominence. The outer posterior prominence reaches the same level posteriorly as the inner posterior prominence ( Fig. 9 View Figure 9 C, G). The bulla bears a deep interprominential notch posteroventrally.
The anterior margin of the tympanic is pointed and bears an incipient anterior spine ( Fig. 9 View Figure 9 A, E). The ventral margin of the tympanic rises anterodorsally ( Fig. 9 View Figure 9 B, F). The anteriormost margin of the tympanic, corresponding to the Eustachian outlet, that dorsally levels with the dorsalmost margin of the involucrum in its posterior region. In all other ziphiids, the dorsalmost margin of the posterior portion of the involucrum is distinctively higher. The dorsal margin of the involucrum is cut by an indentation also visible in medial view. The posterior process of the tympanic is not preserved, but part of the base of the inner posterior pedicle is still present. It is located on the most posteromedial part of the involucrum.
On the ventral surface of the inner posterior prominence, a strong ventral keel is present ( Fig. 9 View Figure 9 C, G). It emphasizes the median furrow separating the two posterior prominences. In ventral view, the lateral margin of the inner posterior prominence is rounded, not flattened as in Ninoziphius platyrostris and Messapicetus gregarius .
A pair of isolated periotics and tympanic bullae discovered in Miocene levels in Kessel (north of Belgium; IRSNB 8243) shares some of the features observed in the holotype of D. mojnum : the anterior margin of the tympanic bearing an incipient spine; the dorsal elevation of the anterior margin of the tympanic; the depression along the anteroventral portion of the pars cochlearis (although not triangular); and the deep anteroexternal sulcus separating the anterior process and the mallear fossa. Without association with other cranial parts, the identification of these isolated earbones remains uncertain.
Stapes. The stapes of the holotype of D. mojnum is still attached to the periotic in the fenestra ovalis. The stapes is conical, and its base is oval. On the posteromedial margin of the stapes, near its head, the muscular process is well developed. At approximately mid-length, the vestigial stapedial foramen is visible. The diameter of the oval head of the stapes corresponds to half the diameter of its base.
Mandible. Measurements of the lower jaw are provided in Table 1 View Table 1 . It is more slender and more elongated than in extant ziphiids, except Mesoplodon grayi ( Fig. 10 View Figure 10 ). The straight symphyseal portion is ventrally semicircular in transverse section and both dentaries are strongly ankylosed. Dorsally, the suture between the two dentaries is attenuated but still discernible. The dorsal surface of the symphyseal portion is medially excavated, particularly on its posteriormost part, forming a U-shape ( Fig. 11 View Figure 11 B).
The alveolar groove is well developed and occupies more than half the length of the mandible. The alveoli are deep, separated by thin interalveolar septae. The alveoli are close to each other along most of the symphysis except at the apex of the mandible. The number of alveoli is estimated to be 29 on each dentary. The original tooth count may be slightly higher, because the posteriormost alveoli are more difficult to discern.
At the apex of the mandibles, in between the anteriormost alveoli, a strong pogonion is present. The anteriormost apical alveolus is the largest one, laterally compressed and anterodorsally orientated. The following subapical alveolus is elliptical, even more elongated anteroposteriorly and laterally compressed ( Fig. 11 View Figure 11 A). On each side of the dentary, two shallower anteroposteriorly elongated alveoli are present posteriorly to the apical and subapical tusk-bearing alveoli. There are no diastemata between the tuskbearing alveoli and the alveoli past them. The rest of them are square-shaped ( Fig. 11 View Figure 11 B).
Two mental foramina are visible on the lateral side of the mandible between the level of the mandibular foramen and the posterior limit of the symphysis. They are elliptical, elongated anteroposteriorly. The most proximal mental foramen is situated at the posterior end of the alveolar groove. Based on the proportions of the skull and comparison with the mandible of Messapicetus gregarius and Ninoziphius platyrostris , at least 20 mm of the mandible is missing posteriorly. Posteriorly to the alveolar groove, the dorsal margin of the mandible is dorsally convex: this area corresponds to a precoronoid crest ( Figs 2 View Figure 2 C, 10B).
Teeth. The number of retrieved teeth for specimen MSM1001x is 75. Most of them can be positioned on the lower jaw. They can be differentiated based on their shape and wear pattern, allowing the distinction of five categories:
1.
The apical tusk ( Fig. 12 View Figure 12 A–C): only one tooth fits in the right apical alveolus. Once in the alveolus, the triangular tooth projects anterodorsally. The root is more developed than the remains of the crown. The margins of the root and the crown are continuous, but can still be distinguished based on presence of enamel on the crown. The crown is almost worn to the root. The surface of wear slopes anteroventrally and slightly lingually toward the oral cavity.
2. The subapical tusk ( Fig. 12 View Figure 12 D–E): this tooth has an elevated root compared to the small crown. The subapical tusk is more conical and less compressed laterally than the apical tusk. The rounded dorsal tip of the crown indicates wear of the tip, not as extreme as for the apical tusk. This tooth is interpreted as a tusk based on its unusual morphology compared to the remaining retrieved teeth. It possesses a much higher root compared to its small crown, a feature shared with the apical tusk (1); the root is compressed transversely and has an oval outline (2); the tusk is anterodorsally directed once fitting in its alveolus (3).
3. The subapical teeth ( Fig. 12 View Figure 12 F–K): there are three teeth in this category. They fit in the two subapical pairs of alveoli posteriorly to the two pairs of apical and subapical tusks. The subapical teeth are flattened transversely, but the root is expanded posteriorly relative to the crown. The root of the subapical teeth is not as developed as that of the tusks. The subapical teeth do have a crown slightly bent medially. They do not exhibit apical wear.
4. The symphyseal and post-symphyseal teeth ( Fig. 12 View Figure 12 L–Q): there are 60 teeth in this category. Their root is squared to rectangular in section, as dorsally high as the crown. Their medially bent crown displays a slightly developed mesial keel. Some of these teeth display an occlusion wear facet extended along the mesial or distal side of the tooth, sometimes extended until the apex ( Fig. 12 View Figure 12 V, W). Others also display a slightly trun- cated apex due to apical wear. Several of the teeth described above may be part of the maxillary tooth row as well.
5. The proximal teeth ( Fig. 12 View Figure 12 R–U): these two teeth are similar in shape to the subapical teeth but differ in the well-developed occlusion facets present from the apex to the mesial or distal side of the tooth ( Fig. 12 View Figure 12 X). It can be explained by the position: situated more posteriorly, they are subject to interlocking as with symphyseal/post-symphyseal teeth. The teeth could also be positioned in the maxilla.
The 16 other teeth are too fragmented to be placed along the alveolar grooves.
Postcranial remains
Hyoid apparatus. The basihyal is subrectangular. It is 8 mm thick medially, and progressively thickens toward the lateral articulation with the thyrohyal to a maximum of 18 mm ( Fig. 13 View Figure 13 ). Additional measurements are provided in Table 5 View Table 5 . The dorsal surface is smoothly longitudinally concave unlike its flatter ventral surface. The articular processes are short and anteriorly projected. The surface of articulation with the thyrohyal has an oval outline marked by small foramina. The roughly complete left thyrohyal is 96 mm long, 25 mm wide and 16 mm thick ( Fig. 13 View Figure 13 ). In dorsal view, it is rectangular, curves dorsally and tapers posteriorly. It is particularly thickened at the level of the articulation with the basihyal and at mid-length.
Atlas. The atlas was subject to anteroposterior compression resulting in deformation of the articular facets for the occipital condyles ( Fig. 14 View Figure 14 A, B). The posterior face of the atlas is difficult to interpret; no articulation facet with the axis is distinguished. Any degree of fusion between the atlas and axis cannot be assessed. The atlas is 158 mm wide and estimated to be 38 mm thick. The articular facets for the occipital condyles are still preserved. They have a crescent shape with rectilinear medial margins, originally forming a V. They are estimated to be at least 43 mm wide and 57 mm high each. Between the articular facets, a bony bridge joins them and forms the ventral margin of the neural canal. The bridge is ventrally marked by a mediolaterally elongated fossa ( Fig. 14 View Figure 14 A). The neural canal was originally dorsoventrally compressed, and dorsally margined by the partially preserved neural arch, but the shape of the neural spine remains unknown. Its thickness cannot be assessed, but the spine was orientated posterodorsally. The lower transverse process of the atlas is thick and well individualized from the main body of the vertebra, curved posteriorly. A rounded and thick medial process is visible posteroventrally, with a triangular outline.
Axis. The axis is less well preserved than the atlas, with the left and the ventral parts missing. The anterior face is too damaged to be interpreted. Posteriorly, the axis is still articulated with C3, but these two cervicals are not fused. This interpretation is based on a part of the anterior articulation facet of the centrum of C3, which is still visible, and because C3 is shifted to the left from the axis ( Fig. 14 View Figure 14 E). The right lower transverse process of the axis is partially preserved, but has been displaced more ventrally.
Other cervical vertebrae. Three other cervical vertebrae are preserved: C3, C5 and C6. Measurements are provided in Table 3 View Table 3 . None of these cervical vertebrae is fused.
C3 is still attached to the axis ( Fig. 14 View Figure 14 C–E). Its posterior articular facet is transversely wider than dorsoventrally high. The outline of the pedicles is still visible and a piece of the right lower transverse process can still be distinguished ventrolaterally.
The two last cervical vertebrae are both represented by a subcomplete centrum. They are interpreted as C5 and C6 based on the lack of posterior articular facets for the first pair of ribs (characteristic of a C7) and the presence of a large transverse foramen on the transverse process, absent on C3 and C4 in extant ziphiids and Ninoziphius platyrostris (de Muizon, 1984) . The two vertebrae are perfectly matching, their lower transverse processes following each other. The centrum of both the vertebrae is rectangular in outline. Ventrally, both of them exhibit two excavations separated by a longitudinal keel. The pedicles forming the neural arch are compressed anteroposteriorly, but only the basis is partially preserved. The lower transverse processes are situated on the ventrolateral margins. On the C5, the left lower process is complete. It is almost straight, anterolaterally orientated, and is 16 mm thick anteroposteriorly and 37 mm long.
Thoracic vertebrae. Two thoracic vertebrae can be identified, but their exact position is unknown ( Fig. 15 View Figure 15 ).
Vertebra A is interpreted as a thoracic between 1 and 3 ( Fig. 15 View Figure 15 A–C). Judging from the size of the posterior articular facets of thoracic A, it could be a T1, but without the whole thoracic series it remains uncertain. The centrum is dorsoventrally compressed, larger than for the last cervical vertebra. Two posterior articular facets for ribs are situated along more than half the posterolateral margins of the centrum. Size and width of the pedicles are similar to those of the previous vertebrae. The neural canal was originally triangular.
The thoracic vertebra B is the best preserved with a partial neural spine and neural canal. Its body is transversely wider than dorsoventrally high, a common feature of the first thoracic vertebrae. Thoracic B is interpreted as a thoracic vertebra post-T4. Posterior articular facets for the ribs can be observed along the dorsolateral margins of the vertebra. Their position is similar to the 4th and 5th thoracic vertebrae of Mesoplodon grayi (specimen MNHN 1877- 329). The body of the vertebra is 48 mm long anteroposteriorly. The lateral flanks are concave. The right transverse process is better preserved. It is robust, bearing a deep articular facet for a rib. Anteriorly, an elongated articular facet on the prezygapophysis is indicative of a strong postzygapophysis on the preceding vertebra. The prezygapophysis in itself is robust. The dorsal roof of the neural arch is partially preserved. The neural arch was triangular and bore a posterodorsally orientated neural spine. The total length of the neural spine is not assessable but its maximum anteroposterior thickness is 29 mm.
Ribs. The ribs are mostly shattered and fragments have been displaced ( Fig. 16 View Figure 16 ). They are tentatively identified and paired based on size and shape.
The pair of ribs 1 is partially preserved ( Fig. 16 View Figure 16 A, B). The left rib is the most complete. It is fractured; the head and the angle are not preserved. It can be identified by its large and flattened body. It is the widest rib with a maximum body width of 56 mm.
Pair 2 is also partially preserved ( Fig. 16 View Figure 16 C, D). Right rib 2 is only represented by a thickened angle matching in size and shape with left rib 2.
Rib 3 is the most complete ( Fig. 16 View Figure 16 E). It was originally flat, with a maximum lateromedial width of 46 mm and more than 400 mm long dorsoventrally.














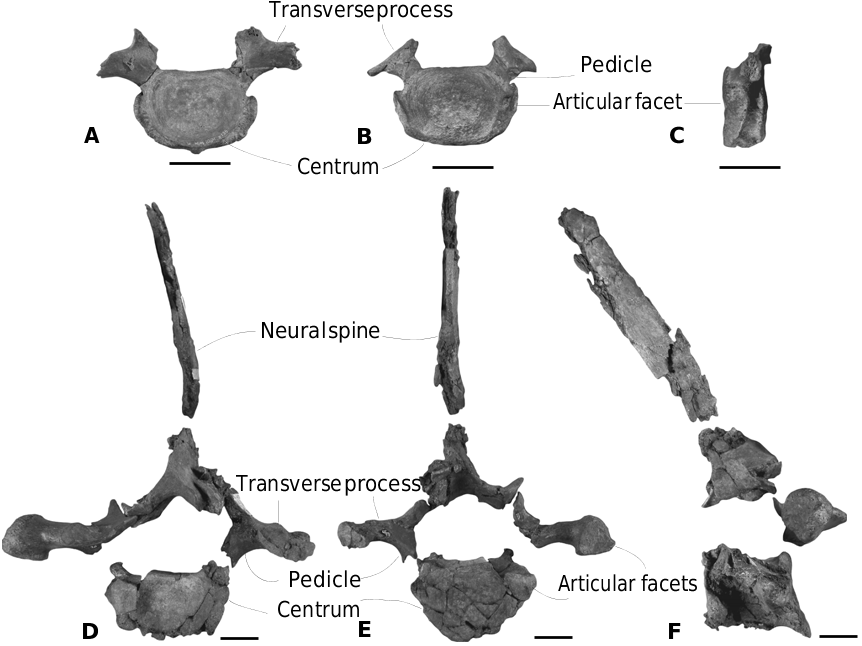

Table 1. Measurements of the skull and mandible of MSM 1001 x (holotype), Dagonodum mojnum gen. nov., sp. nov.
| Feature | MSM1001x |
|---|---|
| Palatal trough anteroposterior length | 324 |
| Palatal trough maximum transverse width | 41 |
| Preserved distal part of the rostrum | 64 |
| at mid-length transverse width | |
| Left premaxillary sac fossa transverse width | 45 |
| Premaxillary sac fossae transverse width | 131 |
| Left ascending process of premaxilla | 32 |
| minimum transverse width | |
| Left premaxillary crest transverse width Left nasal anteroposterior length | 44 49 |
| Left nasal transverse width | 36 |
| Medial suture of nasals anteroposterior | 49 |
| length | |
| Joined nasals maximum transverse width Brain case maximum transverse width | 92e 204 + |
| Brain case maximum anteroposterior length | 124 + |
| Mandibular alveolar groove | 631 |
| anteroposterior length | |
| Symphyseal portion anteroposterior length | 356 |
| Symphyseal portion posterior to | 84 |
| apical alveolus transverse width |
Measurements in mm. e, estimated; +, nearly complete.
Table 2. Measurements of the periotic and the tympanic bone of MSM 1001 x (holotype), Dagonodum mojnum gen. nov., sp. nov.
| Feature | MSM1001x |
|---|---|
| Periotic maximum anteroposterior length | 35 |
| Periotic maximum transverse width | 24 |
| Pars cochlearis on the periotic maximum | 20 |
| anteroposterior length | |
| Pars cochlearis on the periotic maximum | 14 |
| transverse width | |
| Anterior process on the periotic maximum | 14 |
| anteroposterior length | |
| Anterior process on the periotic maximum | 9 |
| transverse width | |
| Posterior process on the periotic maximum | 14 |
| anteroposterior length | |
| Posterior process on the periotic maximum | 10 |
| transverse width | |
| Lateral tuberosity in lateral view on the | 8 |
| periotic maximum transverse width | |
| Tympanic maximum anteroposterior length | 38 |
| Tympanic maximum transverse width | 22 |
| Inner posterior prominence of the tympanic | 10 |
| maximum transverse width | |
| Outer posterior prominence of the tympanic | 12 |
| maximum transverse width | |
| Tympanic maximum dorsoventral height in | 15 |
| lateral view as preserved | |
| Involucrum indentation on the tympanic | 12 |
| dorsoventral height in medial view |
Measurements in mm and taken in ventral view unless
noted otherwise.
Table 3. Measurements of the cervical and thoracic vertebrae of MSM 1001 x (holotype), Dagonodum mojnum gen. nov., sp. nov.
| Feature | Atlas | C3 | C5 | C6 | A | B |
|---|---|---|---|---|---|---|
| Total lateromedial width | 158 | – | – | – | – | – |
| Centrum lateromedial | 48e | – | 62e | 65 | 73 | 93 |
| anterior width | ||||||
| Centrum dorsoventral | 57e | – | 72e | 70 | 60 | 51 |
| anterior height | ||||||
| Centrum lateromedial | – | 66e | 78 | 73 + | 87 | 83e |
| posterior width | ||||||
| (without articular | ||||||
| facets) | ||||||
| Centrum dorsoventral | – | 54e | 71e | 66 + | 63 | 46 |
| posterior height | ||||||
| Centrum | 38e | 28 | 22 | 28 | 29 | 56 |
| anteroposterior length |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.