
Arambourgthurus, Tyler, 2000
|
publication ID |
https://doi.org/10.5281/zenodo.5371235 |
|
persistent identifier |
https://treatment.plazi.org/id/03AC87C7-7D4E-4820-F82F-405FFE5B7494 |
|
treatment provided by |
Marcus |
|
scientific name |
Arambourgthurus |
| status |
gen. nov. |
Genus Arambourgthurus n. gen.
TYPE SPECIES. — Naseus scombrurus Arambourg, 1956 . ETYMOLOGY. — For Camille Arambourg, the distinguished paleontologist whose publications and arduous field work in many parts of the world greatly enriched our knowledge of Tertiary fossil fishes and the collections of the Muséum national d’Histoire naturelle, Paris. Prof. Arambourg first described the type species of the new genus and noted one of its most interesting and unique features. And “ thurus ”, a common ending for generic names among acanthurids.
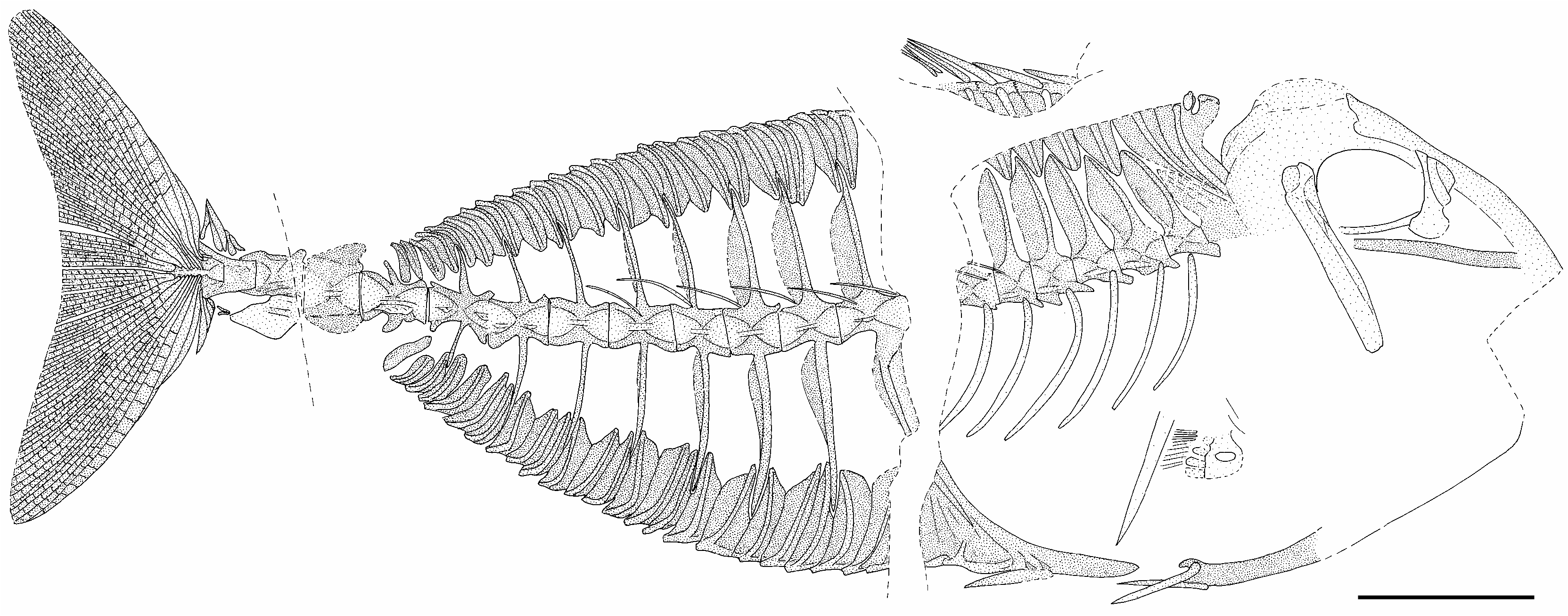
DIAGNOSIS. — Unique among all acanthurids, both RE- DESCRIPTION fossil and Recent, by two derived features so: extensive over- Most of the bones of the head are poorly prehypurostegy, with the caudal-fin rays broadly lapping the hypural plate that their bases are separated served and somewhat dislocated, and little of by only a narrow horizontal space along the midline of interest can be noted; no teeth are preserved in the plate ( versus hypurostegy absent or only very mod- either specimen (the edges of the jaws are lost in erately developed spaces vacant); the versus first, second, and third the paratype and undistinguishable in the holointerneural (only the second and third, or only the third, or none vacant). type). The hyomandibular is well-preserved and clearly exposed in the paratype; it has a relatively flat surface, without a horizontal crest along its
Arambourgthurus scombrurus upper lateral region below its articulation with
( Arambourg, 1956) the skull. The frontal, lateral ethmoid, ethmoid,
( Figs 1-5 View FIG View FIG View FIG View FIG ) and parasphenoid below and in front of the eye are clear in the paratype, and of normal acan-
MATERIAL EXAMINED. — Holotype, Muséum national thurid condition. d’Histoire naturelle to (MNHN. mm) 1939 -6-5 ( EIP 5), single There are six dorsal-fin spines. The first spine is a plate, head right, c 85 SL; the bones of the head are much displaced, and the last (urostylar) cen- short, non-protruding, cap of bone in both specitrum and caudal fin are missing, the vertebral column mens, the remains of which are present over the ending th with only an impression of the penultimate median flange arising from the dorsal surface of (12) centrum and its posteriorly elongate neural and haemal spines. Paratype, MNHN 1939- 6-7 g (EIP 7g) the first dorsal-fin pterygiophore, around which and 7d (EIP7d), 146mm SL; 7g is a relatively com- the cap-like spine would rotate in a locking plete specimen but 7d is only fragmentary; however, mechanism. The other dorsal-fin spines are best 7d has the best preserved remains of the bases of the preserved in the holotype: the second spine is hypurostegic caudal-fin rays.
prominent and slightly longer than the third and
TYPE LOCALITY AND HORIZON. — Both specimens are fourth spines; the fifth spine is missing; the sixth from the Istehbanat locality of Iran, of lower spine is the longest in the series, being about one- Oligocene age (Rupelian), about 35 My. third longer than the second spine. The first two DIAGNOSIS. — That of the new genus, of which it is dorsal spines are borne in supernumerary associapresently the only known representative. tion on the first pterygiophore. In the paratype ,
Tyler J. C.
the second and third dorsal spines are missing, but the distal end of the fourth spine and all of the fifth and sixth spines are present above a fracture through the pterygiophores. There is no evidence of any kind of posterolateral shielding flange from the first pterygiophore around the cap-like first spine. The dorsal-fin rays are fragmented and scattered, but there are about 24-26 pterygiophores behind those of the spiny dorsal fin and therefore about that many rays. There is no supraneural (predorsal) bone between the first dorsal pterygiophore and the skull.
The series of anal-fin spines are incomplete in both specimens. In the holotype, the third spine is fully preserved, situated in about the middle of the ventral surface of the second anal-fin pterygiophore. Only the distal end of the second anal spine is preserved in the holotype; the base of the spine and the region immediately in front of it where the first spine would be is missing. The base of the second spine is situated toward the posterior end of the ventral surface of a forward extension of the first anal-fin pterygiophore, but this extension is not well-preserved. In the paratype, the third anal spine is missing but the second spine is completely preserved, and immediately in front of its base is a small cap of bone that is either the short, non-protruding, cap-like first anal spine or the rounded median flange of the pterygiophore around which the first spine would rotate in a locking mechanism. The bases of the first and second anal spines are situated just below lateral strengthening ridges toward the posterior end of the first anal-fin pterygiophore, and this pterygiophore has a long forward extension to a level in front of the ventral end of the postcleithrum. There is no evidence of any kind of posterolateral shielding flange from the first anal pterygiophore around the cap-like first spine. The anal-fin rays are fragmented and scattered, but there are about 24 pterygiophores in both specimens behind the two for the anal spines and therefore about that many rays.
In the pectoral-fin girdle, the scapular foramen is complete and the postcleithrum is a single bone. About 13-14 pectoral-fin rays are evident in the paratype.
New genus of hypurostegic Oligocene surgeonfish
The pelvic fin has a stout spine; in the holotype there are either two or three rays internal to each spine, and in the paratype there seem to be two rays associated with each spine. It is clear that there are no more than three rays in each pelvic fin. The posterior process of the pelvis is short. The pelvis becomes relatively deep anteriorly at its articulation between the cleithra; the ratio of the length of the pelvis to its depth at the posteri- or edge of the cleithrum is 13%. There does not seem to be any significant anteroventral prong (subpelvic keel) from the lower anterior region of the pelvis below the cleithrum.
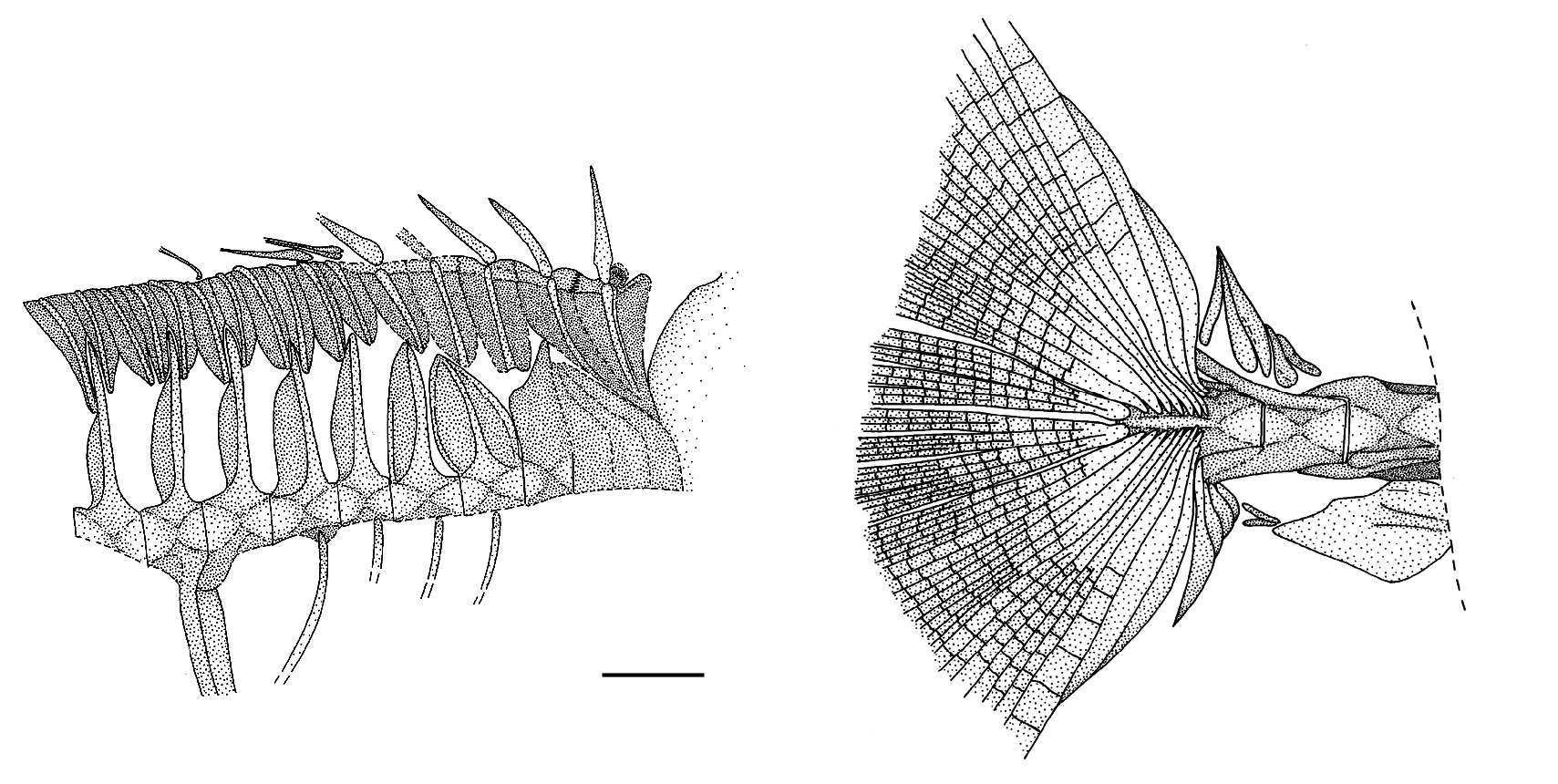
There are 16 principal caudal-fin rays and seven procurrent rays above and below; the innermost two principal caudal-fin rays, one from the upper lobe and one from the lower lobe, are deeply divided to the base (Arambourg interpreted these as four rays, accounting for his statement of 18 principal rays). Above and below the two innermost rays, the other principal rays more anteriorly increasingly overlap the hypural plate, and the proximal ends of the rays from the upper and lower lobes are only separat- ed by a narrow space along the middle region of the plate, which middle region is slightly upraised as a crest. The hypural plate bears shallow grooves (visible in EIP 7g because the fractured bases of the rays have adhered to EIP7d) to accommodate the overlapping slender proximal ends of the rays.
There are 13 caudal vertebrae, as seen especially clearly in the paratype. The paratype has a fracture just in front of the centrum of the first caudal vertebra; this fracture continues into the middle region of the long haemal spine of the first caudal vertebra and of the dorsal shaft of the first anal-fin pterygiophore that this haemal spine supports along its anterior edge. The neural and haemal spines of the first to ninth caudal vertebrae are of decreasing length and support soft dorsal- and anal-fin pterygiophores. The neural and haemal spines of the tenth and eleventh caudal vertebrae are low but broad, whereas those of the twelfth caudal vertebra are elongated posteriorly above and below the urostylar thirteenth centrum. As exposed on EIP 7g, it is clear that hypurals 1-4 are fully fused together into a single plate; hypural 5 cannot be distinguished as a separate element, perhaps because of the ray-accommodating grooves along the upper part as well as elsewhere on the fused plate. Most of the caudal vertebrae have prominent low crests for muscle attachment horizontally along the middle of the centra. There are two epurals and a small, short uroneural just below the bases of the epurals. There is a stout parhypural closely applied to the lower edge of the urostylar centrum and hypural plate. The least depth estimated for the fleshy caudal peduncle, at the level of the eleventh caudal vertebra, is between 4.3-4.7% SL.
The six more posterior of the abdominal vertebrae are relatively well-preserved and individually distinct, but those more anteriorly are poorly preserved, fractured, and not individually distinct. Given that Arambourgthurus has 13 vertebrae in the caudal series, like all other acanthurids, it is reasonable to presume that there were nine vertebrae in the even more conservative abdominal series, like all other acanthurids; moreover, there is space between the rear of the skull and the front of the clearly preserved sixth from last abdominal vertebra for about three more vertebrae.
The ventral shafts of the first two dorsal-fin pterygiophores are strongly curved anteriorly toward the lower region of the rear of the skull, and their lateral strengthening ridges converge just above the level of the centra and neural canal, with no neural spine in-between the two shafts. The neural spines of the first three abdominal vertebrae are closely applied to one another and squeezed in-between the rear of the strengthening ridge of the second dorsal-fin pterygiophore and the front of the fourth neural spine.
The fourth and more posterior abdominal vertebrae in the paratype have well-developed ventral prezygapophyses; these are less clearly preserved in the holotype. Sturdy pleural ribs clearly are present on the fourth to ninth abdominal vertebrae, but whether pleurals were present more anteriorly is unclear. Slender epineurals are present from at least the fifth abdominal to the fifth caudal vertebrae.
In the paratype, there is a single dorsal pterygiophore between the neural spines of the fourth and fifth abdominal vertebrae but two pterygiophores here in the holotype. The next two interneural spaces (behind the fifth and sixth neural spines) have single pterygiophores present, posterior to which the interneural spaces have either one or two pterygiophores, or, far posteriorly, three pterygiophores.
The body is covered with minute scales bearing a central upright spinule or a tiny cluster of spinules, giving a shagreen-like appearance to the surface, with the limits of the individual scales not easily discernable. The lateral line is clear along the middle region of much of the upper half of the body.
In the paratype there is a large ( 8.5 mm) caudal peduncular fixed-plate scale with a horizontal crest. This fixed plate is displaced ventrally below the eleventh ( PU3 ) and twelfth ( PU2 ) caudal centra. In the holotype, the single fixed plates from both sides are displaced just above and just below PU3 ; as in the paratype, each fixed plate has a horizontal crest.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.