
Monstrillopsis chathamensis, Suárez-Morales & Morales-Ramírez, 2009
|
publication ID |
https://doi.org/ 10.1080/00222930902894419 |
|
publication LSID |
lsid:zoobank.org:pub:2EB343F4-0486-4096-B53E-C650357406B0 |
|
persistent identifier |
https://treatment.plazi.org/id/03ADD769-D428-816A-ECB6-FA5754D9CD80 |
|
treatment provided by |
Felipe |
|
scientific name |
Monstrillopsis chathamensis |
| status |
sp. nov. |
Monstrillopsis chathamensis sp. nov.
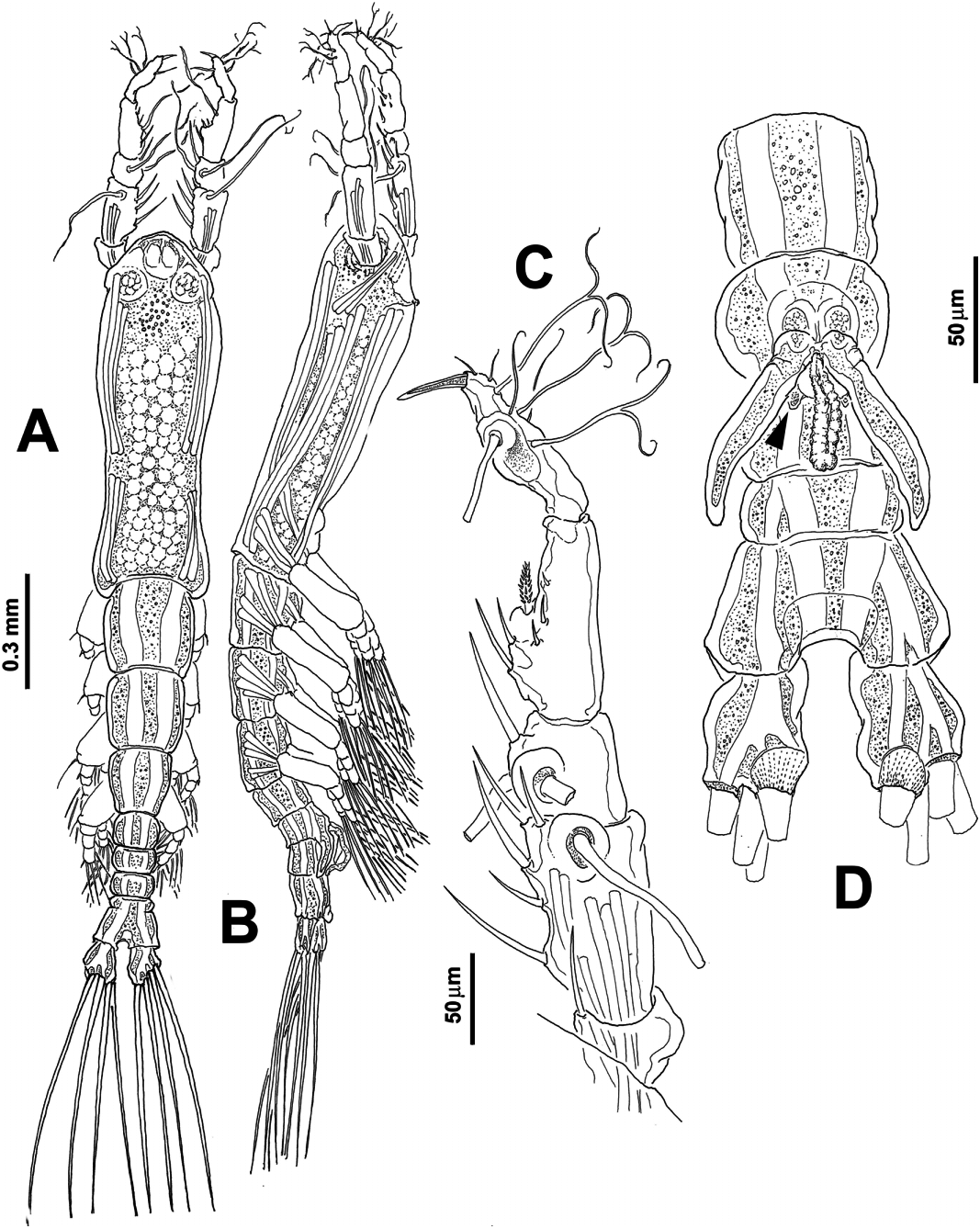
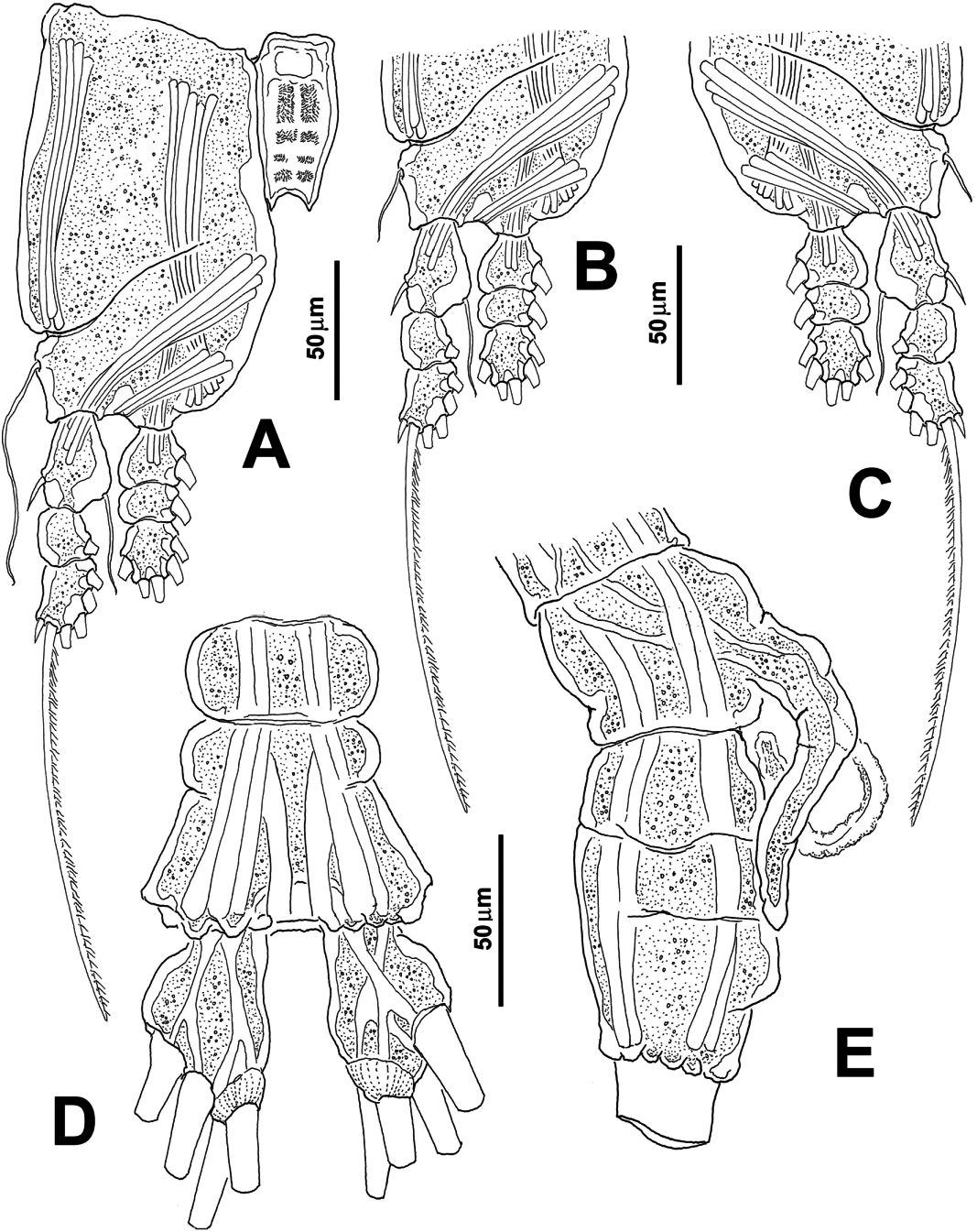
( Figures 4 View Figure 4 , 5 View Figure 5 )
Material examined
Holotype. Adult male, dissected, permanent slide mounted in glycerine sealed with Entellan. Plankton collection, Chatham Bay, Isla del Coco, Costa Rica (5°33′ 01.58′′ N, 87°02′ 18.57′′ W). Specimen collected 12 November, 2000, by J. Picado, semi-permanent slide (ECO-CHZ-03958). GoogleMaps
Paratypes. Four adult males, same date and locality, undissected, ethanol-preserved (ECO-CHZ-03959).
Description
Body length of adult male holotype: 0.93 mm, measured in dorsal view from anterior end of cephalothorax to posterior edge of anal somite, caudal rami excluded. Mean length of specimens: 0.93 mm, range: 0.90–0.95 mm. Cephalothorax incorporating first pedigerous somite representing about 51% of total body length ( Figure 4A,B View Figure 4 ). Body slender, anterior half of cephalothorax wider than posterior half. Middle part of cephalothorax narrow, gradually expanding posteriorly. Dorsal ocelli present, pigment cups relatively small, separated by the distance of about two-thords of the ocellus diameter, poorly developed, weakly pigmented, rounded in dorsal view. Ventrally, anterior cephalic surface with reduced, faint ornamentation represented by a few transverse striations between oral papilla and antennule bases. Oral papilla reduced, weakly protuberant, located about 0.11 of way back along ventral surface of cephalothorax ( Figure 4B View Figure 4 ).
Antennules moderately long, with five well-defined segments; antennular length 0.33 mm, representing close to 30% of total body length, and about 61% as long as cephalothorax. Length ratio of antennular segments, from the proximal to distal being: 8: 28: 12: 29: 23 (= 100) ( Figure 4C View Figure 4 ). Segments armed with 0-I; 1-V; 2-I; 1-V; 5-II+aes setae (in Arabic numbers), spines (in Roman numbers), and aesthetascs (aes). Distal antennular segment geniculate, with three subequal, distally dichotomously branched setae aligned near outer distal end. As usual in male Monstrillopsis , terminal antennular segment modified, with inner margin rounded, moderately expanded and distal half forming sabre-like structure with elongated distal element. In terms of the setal nomenclature of Grygier and Ohtsuka (1995) for female monstrilloid copepod antennules, elements in first (1), second (2d 1,2, 2v 1–3, IId), and third (3, IIIv, IIId) segments complete. Fourth with elements 4v 1–3, 4d 1,2. In terms of the setal nomenclature proposed by Huys et al. (2007) for the armature of the distal segment of male monstrilloids, elements 1–5 present on anterior margin, dichotomously branched elements A–C on posterior margin ( Figure 4C View Figure 4 ). Distal segment with swollen inner margin and with elongate distal element.
First pedigerous thoracic somite incorporated into cephalothorax. This and the succeeding three pedigers each bearing well-developed swimming legs, all with three-segmented endopods and exopods and with same armament pattern, except for leg 1 exopod which has one fewer seta on distal segment ( Figure 5A–C View Figure 5 ). Coxa of each pair unarmed, joined by subrectangular intercoxal sclerite. Sclerites with anterior surface covered by fields of minute spinules ( Figure 5A View Figure 5 ). Basis separated from coxa by diagonal articulation in legs 1–4. Basipodal seta of third swimming leg 3.5 times longer than in other legs. Armature of swimming legs is shown in Table 2:
Fifth leg absent, fifth pedigerous somite globose. Succeeding somite (genital somite) short, lateral margins rounded; genital somite with genital complex, represented by ventrally protruding genital apparatus. Apparatus with two divergent lappets arising from middle of somite ( Figure 4D View Figure 4 ). In lateral view, shaft curved downwards, distal half of lappets bent posteriorly, reaching proximal margin of anal somite ( Figure 5E View Figure 5 ). Base of arms with small rounded processes, one on each branch (arrowed in Figure 4D View Figure 4 ); most specimens with spermatophores partially ejected, protruding between genital lappets ( Figure 4D View Figure 4 ).
As usual in male Monstrillopsis , urosome consisting of five somites: fifth pedigerous somite, genital somite (with genital complex/apparatus) and three free somites. In dorsal view anal somite longest, partially fused with preceding somite, suture visible in ventral view, but only partially in dorsal position; anal somite with roughly corrugated dorsal posterior margin ( Figure 5D View Figure 5 ). Ratio of lengths of genital somite and three succeeding free somites: 36: 19: 16: 29 (= 100).
Caudal rami subrectangular, about 1.4 times longer than wide. Caudal rami each armed with four biserially setulated setae ( Figures 4D View Figure 4 , 5D View Figure 5 ). All caudal setae subequal in length and width.
Type locality
Chatham Bay , northern coast of Isla del Coco (5°33′01.58′′ N, 87°02′18.57′′ W), Costa Rica GoogleMaps .
Etymology
The species name refers to the type locality, Isla del Coco , Costa Rica. Remarks
The male specimens from Isla del Coco are assigned to the genus Monstrillopsis on account of their having the main characters of males of this genus, including three free postgenital somites, four setae on each caudal ramus, and a modified distal antennular segment (see Suárez-Morales et al. 2006, 2008). In their revision of Monstrillopsis, Suárez-Morales et al. (2006) did not include a definition of the males. One of the main generic characters of the females of Monstrillopsis is the presence of four caudal setae; however, this character is not equally valuable in the males, because males of some species of Cymbasoma have four caudal setae ( Suárez-Morales 2007). The urosomal segmentation with three postgenital somites and the antennule morphology with a modified distal segment, provide the genus-defining characters ( Huys and Boxshall 1991; Suárez-Morales et al. 2006). Males are known for only a few species of this genus ( Suárez-Morales et al. 2006): M. fosshageni Suárez- Morales and Dias, 2001, M. sarsi Isaac, 1975 , M. dubioides Suárez-Morales, 2004 , Monstrillopsis sp. of Huys and Boxshall (1991), and M. chilensis Suárez-Morales, Ramírez and Derisio, 2008 . The new species differs from the other known species of Monstrillopsis in the structure of the genital complex. In M. sarsi the genital lappets are widely divergent with truncate tips, and they are long, reaching the middle of the anal somite, whereas in the new species the lappets are not divergent and are relatively shorter, reaching only to the anterior margin of the anal somite, while their tips are distally attenuated. In M. chilensis the genital lappets resemble those of the new species, but they are relatively longer than in C. guerrerense and their margins are roughly corrugate compared with a smooth surface in the new species. In M. dubioides , M. fosshageni and Monstrillopsis sp. , the genital lappets have a quite different structure, being fused to form a single shaft with distally bifurcate rounded expansions ( Sars 1921; Suárez-Morales and Dias 2001; Huys and Boxshall 1991).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |