
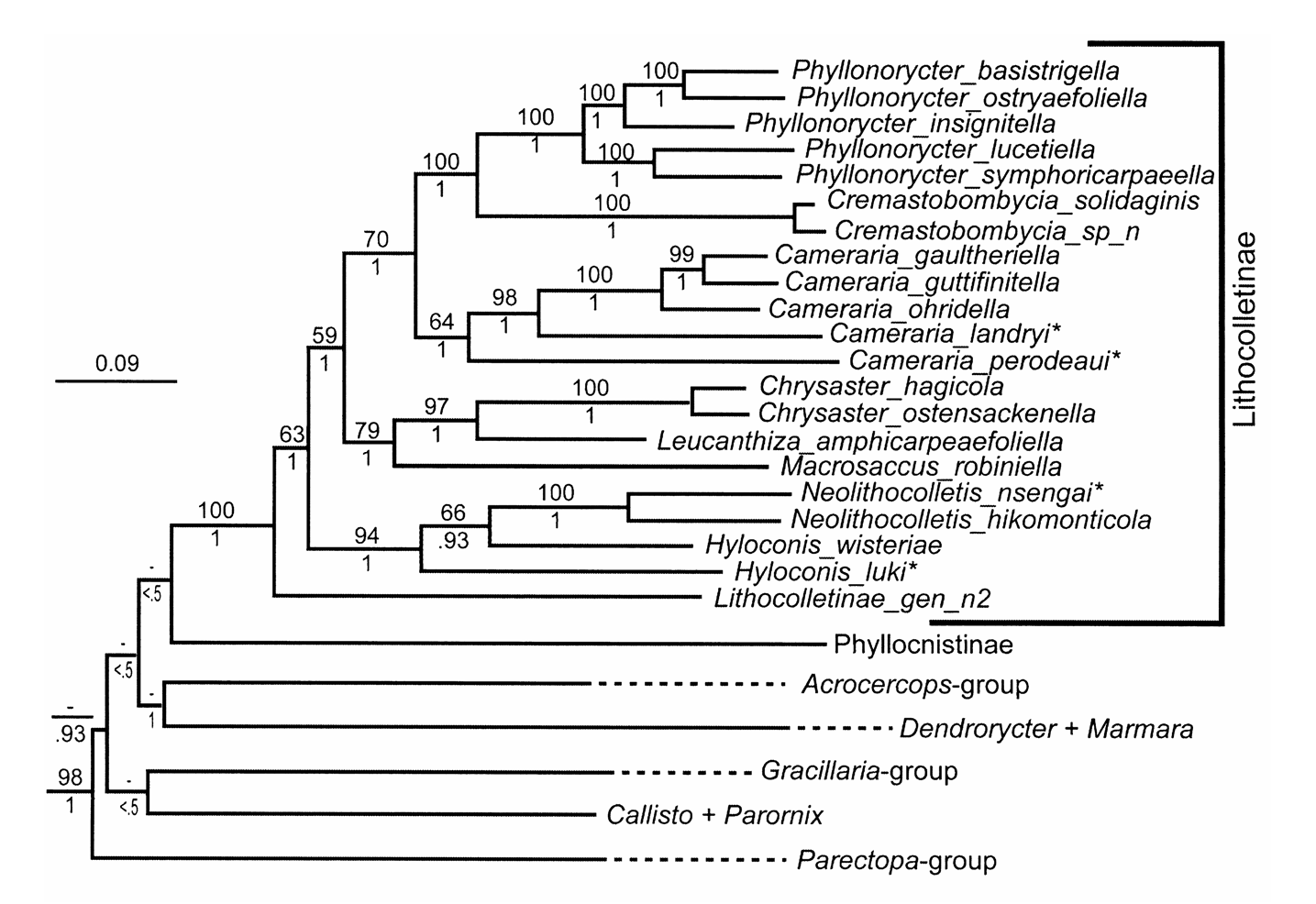
Neolithocolletis Kumata, 1963
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3594.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:B00799F3-F397-438C-B1E1-A8440E636921 |
|
DOI |
https://doi.org/10.5281/zenodo.6308635 |
|
persistent identifier |
https://treatment.plazi.org/id/03ADE350-B108-FF94-F1CF-FD658C9FCCB1 |
|
treatment provided by |
Felipe |
|
scientific name |
Neolithocolletis Kumata, 1963 |
| status |
|
Neolithocolletis Kumata, 1963 View in CoL
Neolithocolletis Kumata 1963: 21–22 View in CoL .
Type species: Neolithocolletis hikomonticola Kumata, 1963 View in CoL , by original designation and monotypy.
Historic account. Kumata (1963) proposed Neolithocolletis to accommodate one Japanese species, N. hikomonticola Kumata, 1963 . Although the latter closely resembles Phyllonorycter , the presence of vein R 2 in the forewing and hairy processes on the sclerotized anellus suggested a new genus was necessary. In the original description of the type species, Kumata (1963: 21) indicated the presence of a setose uncus in Neolithocolletis : “Uncus very elongate, with four apical setae and some microspines on outer surface.” In the following publication Kumata (1993) pointed to an additional character of this genus differentiating it from Cameraria and Hyloconis : the presence of the very wide transtilla. Neolithocolletis remained monobasic until Kumata (1993) added two additional species: a new species, N. kangarensis Kumata, 1993 , and N. pentadesma ( Meyrick, 1919) , the latter of which he transferred from Phyllonorycter . In the same publication, Kumata described three features that distinguish Neolithocolletis from other lithocolletine genera: i) fasciate whitish markings of the forewing margined basally, ii) tegumen of male genitalia with four setae (note: Kumata did not mention an uncus in the additional description of Neolithocolletis ), iii) corpus bursae with many microscopic spine-like signa, and iv) larval characters. Based on the larval characters Kumata (1993: 6) concluded “ Neolithocolletis is very closely related to Hyloconis . ” Our molecular data confirm his hypothesis ( Fig. 4 View FIGURE 4 ). Kumata (1995) discovered a female belonging to Neolithocolletis in the Philippines. Based on N. hikomonticola, Kuznetzov & Baryshnikova (2001) grouped Neolithocolletis with Chrysaster , Hyloconis . and Phyllonorycter . Neolithocolletis hikomonticola was found to be the host of a new Eulophidae (Hymenoptera) species ( Kamijo 1990; Ikeda 1995), whereas N. pentadesma modeled the bionomic study ( Sajap et al. 1996).
To date, three species of Neolitholletis are known from three biogeographical regions: Palaearctic, Oriental, and Afrotropical ( Kumata 1993; Gerlach & Matyot 2006; Bai et al. 2009).
Diagnosis. We define Neolithocolletis as a species-group taxon of the subclade Neolithocolletis nsengai , n. sp. + Neolithocolletis hikomonticola ( Fig. 4 View FIGURE 4 ). The forewing pattern of Neolithocolletis differs from that of Cameraria (except C. fasciata , C. hexalobina , C. loxozona and C. perodeaui ) and Cremastobombycia in that whitish fasciate markings are found on the base of the forewing in Neolithocolletis , except in N. mayumbe . The vertex is tufted as in Cameraria , Cremastobombycia , Hyloconis , Phyllonorycter , and Porphyrosela ; Chrysaster and Protholithocolletis have a smooth vertex. Forewing venation is similar to that of Hyloconis and Protolithocolletis , having an R 2, but differs from Neolithocolletis and Protolithocolletis , lacking a forewing M 2. Hindwing venation is similar to that of Cameraria , Chrysaster , Leucanthiza , and Phyllonorycter , in that it has an unbranched, single M 1, thus differing the genus from Hyloconis , which possesses two hindwing median veins: M 1 and M 2. Male genitalia of Neolithocolletis are symmetrical ( Kumata 1963: 21). The presence of two pairs of apical setae on the tegumen in the male genitalia is unique to Neolithocolletis . The presence of a complete transtilla distinguishes Neolithocolletis from Hyloconis and Cameraria . The corpus bursae of Neolithocolletis bears many microscopic spine-like signulae, whereas other Lithocolletinae genera possesses other types of signa or lack a signum entirely.
Kumata (1993) described diagnostic characters of the larva based on the last instar of N. pentadesma ( Meyrick, 1919) : i) abdomen with a series of subtriangular or elliptical shields both on the dorsal and ventral surface; ii) thoracic legs reduced into a small protuberance with six minute cones on the top; iii) ventral prolegs well developed on third and fourth abdominal segments; and iv) crochets absent on third to fifth abdominal segments and in anal proleg on tenth segment. Chaetotaxy: i) seta XD2 absent on prothorax, ii) seta D2 posterodorsal to D1 on abdomen except on sixth and seventh segments; on those latter segments D2 is posterolateral to D1, far remote from it and thick; iii) D2 on mesothorax and metathorax thickened and set close to D1; iv) lateral group of setae represented by two on all body segments except on eight and nine, L1 longer than L2 in abdominal segments; v) subventral group of setae represented by a single seta SV1 on all abdominal segments and on ventral prolegs; vi) seta V1 absent on sixth to ninth abdominal segments; vii) proprioseptor absent on eighth and ninth abdominal segments; viii) MV3 absent on all abdominal segments. Kumata (1993) indicated that the Lithocolletinae genera could be best diagnosed based on preimaginal stages, especially referring to the last instar. He recognized the following characters as separating Neolithocolletis from other lithocolletine genera: i) Neolithocolletis can be distinguished from Hyloconis by the reduced thoracic legs; ii) in Neolithocolletis seta D2 posterodorsal to D1 on abdomen except on sixth and seventh segments, in Cameraria and Chrysaster the seta D2 is always posterolateral to D1 on all abdominal segments; iii) the ventral prolegs in Neolithocolletis have a single subventral seta and in Cameraria and Chrysaster they have two or three subventral setae; and iv) Neolithocolletis differs from Phyllonorycter by the presence of two lateral setae L1 and L3 on mesothorax and metathorax and L1 and L2 on prothorax and abdominal segments; in Phyllonorycter the lateral group is represented by only one seta L1 on the mesothorax, metathorax and all the abdominal segments. All Neolithocolletis species for which the biology is known feed on Fabaceae ( Kumata 1993) . Mines are irregularly blotch shaped, usually on the disc between two lateral veins of a leaf, mostly on the underside of the leaf, but ocassionally on the upper side as well, whitish, flat, small (less than 2 cm in longest diameter) at maturity, and multiple on the leaflet (up to 40 mines on a single leaflet, occupying the whole lower surface) ( Kumata 1993). Pupation takes place inside the mine within a circular white cocoon that is usually siutated in the center of the mine ( Kumata 1993).
The two species of Neolithocolletis for which we have molecular data, N. hikomonticola and N. nsengai , are grouped together ( Fig. 4 View FIGURE 4 ). Their sequence composition is different from all other taxa we have sampled, and the two species appear to be nested within Hyloconis ( Fig. 4 View FIGURE 4 )
Diagnosis of Afrotropical Neolithocolletis . We assign three Afrotropical species to this genus: the oriental N. pentadesma , discovered in Seychelles ( Gerlach & Matyot 2006), and two native Afrotropical species from central Africa, N. mayumbe De Prins , n. sp., and N. nsengai De Prins , n. sp. Neolithocolletis mayumbe is placed in Neolithocolletis due to: 1) the general similarity of male genitalia to those of N. hikomonticola , N. kangarensis , and N. pentadesma , and 2) because the apex of the tegumen bears two pairs of setae, a diagnostic character of Neolithocolletis ( Kumata 1993) . Neolithocolletis nsengai is placed in Neolithocolletis because of its identical wing venation to that of N. hikomonticola , N. kangarensis , and N. pentadesma (i.e., all possess R 2 in the forewing and lack M 2 in the hindwing). Here we broaden the definition of Neolithocolletis by adding the following morphological characters attained from N. mayumbe and N. nsengai : 1) forewing may have five apical veins (R 2 may be absent), but the tegumen in male genitalia bears two pairs of setae. Four setae on the apex of the tegumen are present in type species, N. hikomonticola , and in N. kangarensis , N. pentadesma , and N. mayumbe ; 2) apex of tegumen in male genitalia may possess one pair of setae, but the forewing has six apical veins (R 2 is present). Six apical veins are present in the type species, N. hikomonticola , and in N. kangarensis , N. pentadesma , and N. nsengai ; 3) the signa area of small spinules on the inner wall of corpus bursae in the female genitalia as in many other Neolithocolletis species can be replaced by a signa line on the caudal part of the corpus bursae, as in N. nsengai . In general, the Afrotropical Neolithocolletis are small moths, having a wing length of ca. 1.6–2.0 mm. The external characters of N. mayumbe and N. nsengai only remotely resemble representatives of the genus from Japan and Malaysia. Wing venation alone cannot serve as a diagnostic character for this genus, and genitalia dissections are necessary. Wing venation of N. nsengai is entirely consistent with Oriental and Palaearctic Neolithocolletis species.
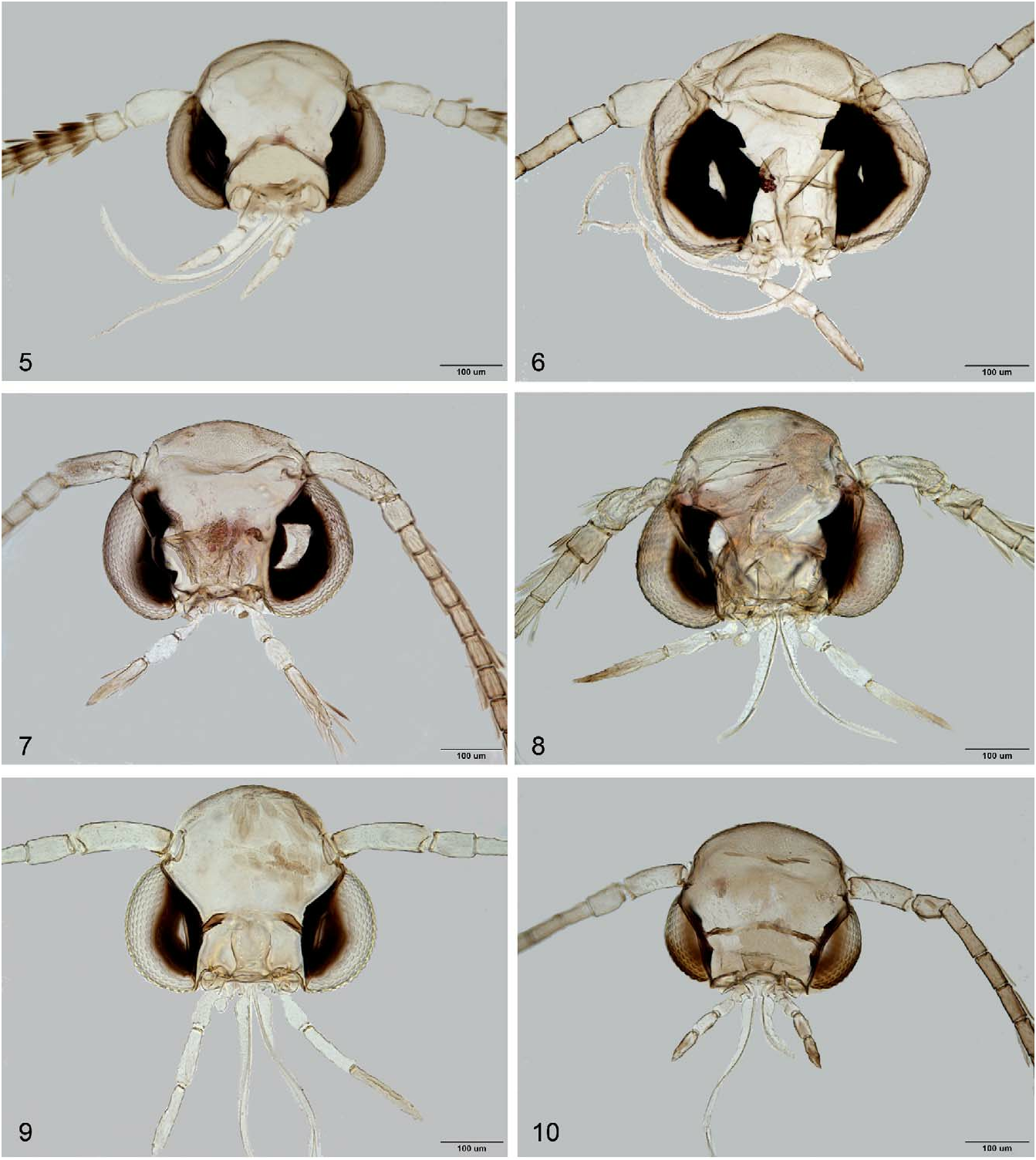
Head: Vertex tufted; frons smooth, white. Maxillary palpus small, rudimentary. Labial palpus short, drooping, filiform, pointed; haustellum moderate in length. Antenna nearly as long as forewing, flagellum thicker than in Phyllonorycter ; pedicel shorter than remaining flagellomeres; scape thicker than flagellomeres, short, with pecten ca. half as long as diameter of compound eye. Vertex plate joined with frontoclypeus, vertex tufted with erected long golden or silver shiny piliform scales, occiput covered with slightly tufted, thick, piliform scales; frons smooth shiny white, consisting of slender pressed piliform scales at central region of frontoclypeus and thicker at lateral sides of frontoclypeus along margin of eye; eyes big, ocular index ca. 0.7, interocular index ca. 1.2. Proboscis developed, naked, ca. 2.0 length of labial palpus. Labial palpus moderate, porrect, filiform, drooping or straight, with ratio of palpomeres from base 1: 1: 1.5 ( Fig. 5 View FIGURES 5–10 ). Maxillary palpus small, rudimental, bi-segmented, apical maxillary palpomere almost globular.
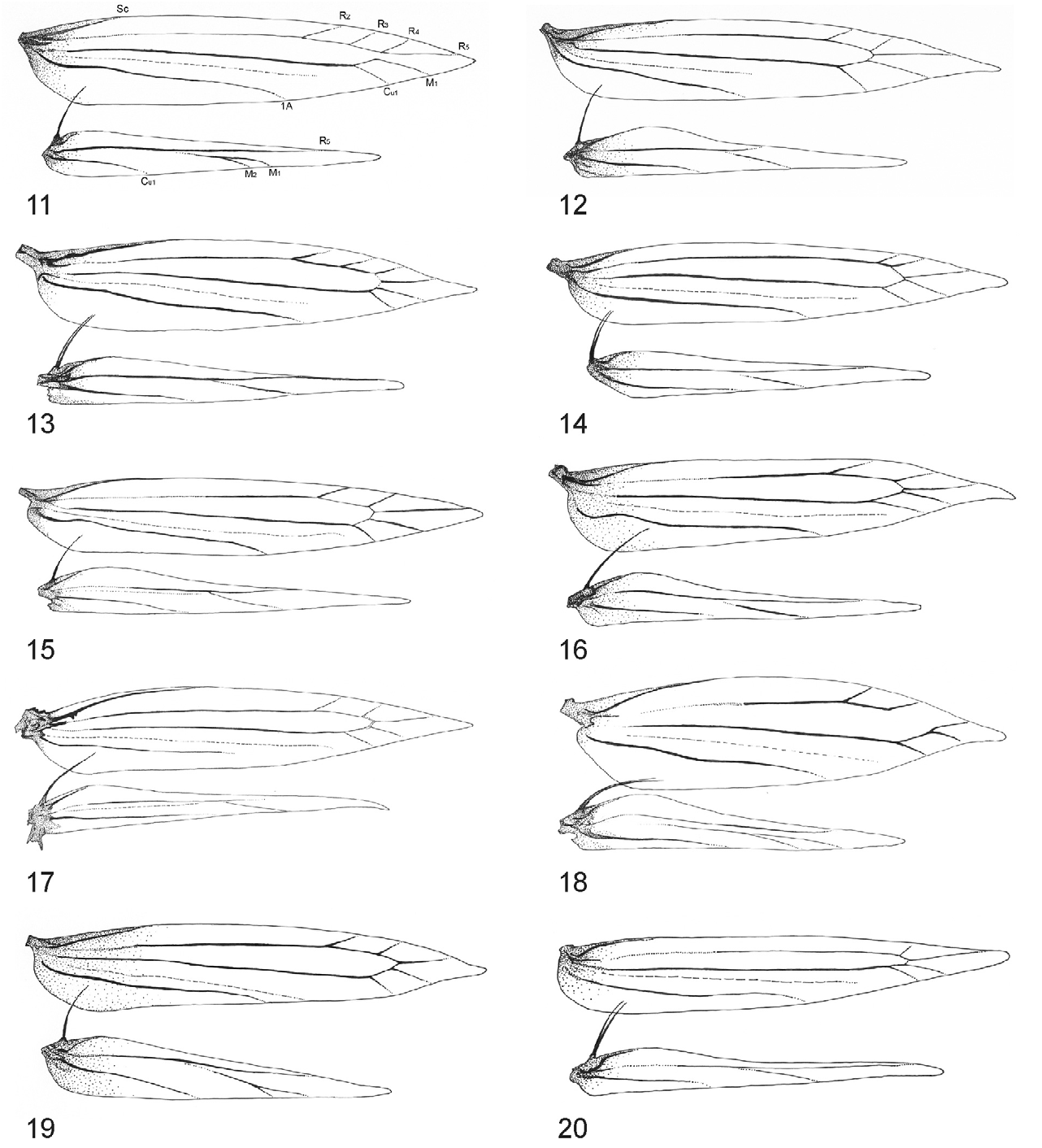
Thorax: Forewing background colour is golden-ochreous, or ferruginous ochreous, background colour shiny golden, or golden ochreous with indistinct fasciate whitish markings, intermixing with the ground colour of forewing and dark beige scales or with very distinct blackish spottish markings; apex of forewing is broadly rounded. Descaled forewing, lanceolate, elongate, and narrowly or shortly pointed: maximum width/length ratio ca. 0.18–0.21. Venation is confined either to 9 veins (in N. nsengai ), apical part, with 6 veins (R 2, R 3, R 4, R 5, M 1, Cu 1); R 2 originates at 5/7 of cell or to 8 veins (in N. mayumbe ) with 5 veins (R 3, R 4, R 5, M 1, Cu 1); M 1 not stalked, the cell between R 4 and M 1 open, R 5 originates from apex of cell to costa, M 1 arises from apex of the cell to termen, Cu 1 separate, R indistinct at basal 1/3, CuP indistinct over entire length, 1A strong, separate. Hindwing lanceolate, ca. 3/4 as long as forewing, with 5 veins: Sc very short, Rs long, running to 3/5 of costa, M 1 single, basal 2/3 of M 1 indistinct, parallel to Rs, Cu 1 strong, simple; A 1 vestigial ( Figs 12, 13 View FIGURES 11–20 ). Retinaculum as a small fold on Sc. Frenulum in male comprised of a single stout bristle; frenula in female comprised of 2 tightly appressed bristles. Legs slender, mid-legs dark ringed, forelegs and hindlegs not ringed; hind tibia with short loosely appressed hairs along apical half, hind tarsus with short appressed hairs on apex of tarsomeres I and II; tarsus ca.1.2× longer than tibia.
Abdomen. Margins of abdominal opening narrowly sclerotized slightly broader towards S 2, the sclerotized margination of abdomen opening well connected on T2 and unconnected on S2; S2 apodemes rather long, ending beyond the opening, very slender, also at base, slender distally. Flap-like sternum VIII small, a little longer than vinculum, subpentagonial (in N. pentadesma ), or gently semi-rounded ( N. nsengai ) or elongated, almost as long as tegumen, spatulate with strongly sclerotized subbasal lateral margins (in N. mayumbe ) either with an angulate apex ( N. pentadesma ) or with gently rouded apex ( N. mayumbe , N. nsengai ).
Male genitalia. Tegumen long, subconical, with very narrow sclerotized arms, weakly sclerotized; apex of tegumen with two long, stiff setae ( N. nsengai ) or four ( N. pentadesma , N. mayumbe ). Valvae symmetrical, narrow, rod shaped ( N. mayumbe ) or slightly enlarged apically ( N. nsengai ), with short sutures or without sutures on ventral surface, with rouded apex bearing thick and short setae ( N. pentadesma ), slender sparse, short setae ( N. nsengai ) or a couple of barbs ( N. mayumbe ). Vinculum crescent-shaped, narrow (in N. mayumbe , N. nsengai ), or slightly broader (in N. pentadesma ) with long slender saccus. Transtilla complete with a pair of small laterocephalic lobes (in N. pentadesma ,), or trapezium-shaped ( N. nsengai ), or narrow bow shaped without laterocephalic lobes ( N. mayumbe ). Aedoeagus is slender tubular, sinuate (in N. pentadesma and N. nsengai ) or straight (in N. mayumbe ). Anellus developed, tubular ( N. nsengai ) or volcano-shaped ( N. pentadesma , N. mayumbe ), juxta either developed in a complex sclerotized structure ( N. pentadesma ) or undeveloped ( N. mayumbe , N. nsengai ).
Female genitalia. Papillae anales slightly compressed laterally, not fused ventrally. Two pairs of apophyses present. Segment VIII rather short in length. Apophyses anteriores initiate at subanterior part of segment VIII. Segment VII separated from segment VIII, with either strongly sclerotized cephalic margin of segment VII ( N. pentadesma ) or with strongly sclerotized arc-shaped scar situated on the entire sternum VII ( N. nsengai ). Ostium bursae opens on sternum between segments VII and VIII. Ductus bursae moderate in length with weakly sclerotized antrum ( N. pentadesma ) or with slight membranization of inner canal ( N. nsengai ). Corpus bursae large pyriform, with many microscopic spine-like signa scattered on inner margin ( N. pentadesma ) or caudal part of corpus bursae is ringed by narrow sclerotized curled line ( N. nsengai ).
Biology. Irregular blotch mine on the abaxial side of the leaf, rarely adaxial, whitish, flat, and small (less than 2 cm at mature stage). Pupation takes place within a circular, white cocoon which is placed inside a round minechamber ( Figs 418, 419 View FIGURES 418–426 ).
Host plant(s). Larvae feed on leaves of Fabaceae .
Distribution. Neolithocolletis mayumbe and N. nsengai occur in the rain forest of Central Africa. Neolithocolletis pentadesma was introduced to Seychelles in 2002 ( Gerlach & Matyot 2006: 23).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Neolithocolletis Kumata, 1963
| Prins, Jurate De & Kawahara, Akito Y. 2012 |
Neolithocolletis
| Kumata, T. 1963: 22 |