
Myosotis antarctica Hook.f. subsp. antarctica, Hook. f., Fl. Antarct.
|
publication ID |
https://doi.org/ 10.1071/SB21031 |
|
DOI |
https://doi.org/10.5281/zenodo.10949728 |
|
persistent identifier |
https://treatment.plazi.org/id/03AE7D0B-FB01-494C-7DCC-FDB4FA57E3C0 |
|
treatment provided by |
Felipe |
|
scientific name |
Myosotis antarctica Hook.f. subsp. antarctica |
| status |
|
Myosotis antarctica Hook.f. subsp. antarctica View in CoL
Myosotis pygmaea Colenso, Trans. & Proc. View in CoL New Zealand Inst. 16: 334 (1884), nom. Illeg. [non Myosotis pygmaea Bertol. View in CoL (in Alessandrini 1840)]. Type citation: ‘Hab. On dry upland open heaths (with Viola perexigua, supra), between Matamau and Danneverke, Waipawa County, 1882–1883: W. C.’ Type: NEW ZEALAND, Southern North Island, heath near Matamau, 2 November 1883, W. Colenso 1609 (lecto [designated by L. B. Moore in H. H. Allan (Ed.), Fl. New Zealand 1: 815 (1961)]: WELT SP004743!); NEW ZEALAND, Southern North Island, nr Matamau, 17 December 1882, W. C. [W. Colenso] (remaining syn: WELT SP004744!).
Myosotis ramificata G.Simpson, Trans. & Proc. Roy. Soc. View in CoL New Zealand 79: 426 (1952). Type citation: ‘ Habitat : Shaded ledges on rocky slopes. Type specimen – in the Herbarium , Plant Research Bureau , Wellington, from sparsely shrub-clad slopes fringing a small flat some three miles south of Luggate , Central Otago.’ Type: NEW ZEALAND, Otago, near Luggate, 28 December 1947, G. Simpson s.n. ( CHR 75720 !) .
Myosotis pygmaea var. drucei L.B.Moore View in CoL in H. H. Allan (Ed.), Fl. New Zealand 1: 816, 973 (1961); Myosotis drucei (L. B. Moore) de Lange & Barkla View in CoL in P. J. de Lange et al. Threat. Pl. New Zealand 438 (2010). Type citation: ‘ Type locality Whanahuia Range , Ruahine Mts. ’ Type: NEW ZEALAND, Southern North Island, Mt Maungamahoe, rocks at summit 5450’, 21 January 1952, J. A. Hay s.n. ( CHR 76820 !)
Typification notes
Moore (1961) cited the type of the name Myosotis pygmaea as ‘W, 4743, W. Colenso’, which is treated here as effective lectotypification. (Note that W was the old herbarium code for what is now WELT.) One other Colenso specimen of M. pygmaea also matches the type citation (WELT SP004744!) and may be original material; however, because it has a different collection date, it is excluded from the lectotype. Material at K mentioned by Moore (1961) was collected earlier and posted to W. J. Hooker in 1848 and 1850, so is not part of the type material (Colenso 1733, K 000787896!; Colenso 2499, K 000787897!: St George 2009, pp. 235, 251).
Description
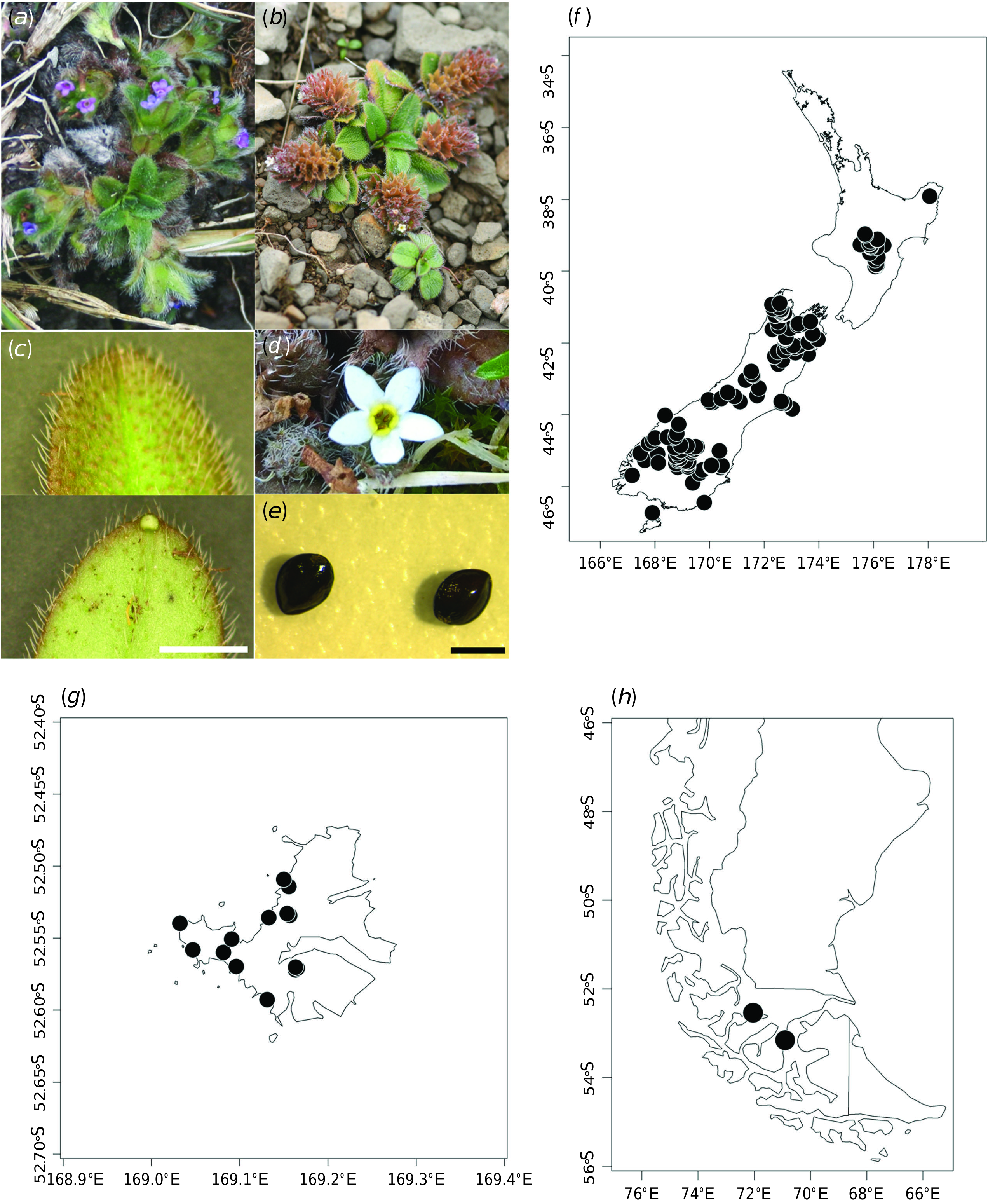
Rosette plants with multiple prostrate branches up to 15(–31) cm long. Rosette leaves 4–22; petioles 1.0–20.0 mm long; lamina margins and apex sometimes curling under, narrowly oblanceolate to very broadly obovate, 3.0–26.0 mm long, 1.5–11.0 mm wide (length:width ratio 1.0–4.0(–6.0):1), bright to dull green to reddish-brown, often with red–brown petioles and mid-veins; apex obtuse, with hydathode on abaxial side; trichomes densely distributed and often overlapping, flexuous, antrorse, patent to erect, spreading at the margins, distributed evenly (on leaf adaxial surface), and sparsely distributed, or on midrib only, or absent (on abaxial surface), (0.2–)0.5–1.1(–2.0) mm long, deciduous with age. Basal cauline leaves not subtending flowers, 1–5 per branch, lamina similar in size and shape to the rosette leaves, with petioles up to 8.8 mm; distal cauline leaves subtending flowers up to 46 per branch, lamina 1.4–16.0(–25) mm long, 0.8–7.0 mm wide, usually sessile. Pedicels up to 1.2 mm long (flowering) or 1.9 mm long (fruiting). Calyx 1.0– 3.5 mm long (flowering) increasing to (2.0–)3.0– 6.5 mm long (fruiting), 1.5–6.0 mm wide at the top at fruiting, lobed to 1/3–3/4 the length of the calyx; with trichomes sometimes of two lengths, longer and antrorse on ribs v. shorter and retrorse in between ribs and near the base (in other instances, the two length classes are not so obvious, and retrorse trichomes are not always present). Corolla (1.0–)1.5–4.0 mm in diameter, white, cream, blue, faucal scales yellow; corolla lobes 0.5–1.5 mm long (0.2–) 0.4–1.1(–1.3) mm wide; corolla tube 0.5–1.2(–1.5) mm wide at faucal scales, 1.2–2.8(–3.3) mm long from base to faucal scales, narrow cylindric. Stamens 5, included; filaments attached below faucal scales, 0–0.3 mm long; anthers 0.3–0.9 mm long, subsessile; style (0.8–) 1.1–2.3 mm long (flowering) to (0.8–)1.1–2.8(–4.8) mm long (fruiting). Nutlets 4, (1.1–) 1.2–1.9 mm long, (0.7–) 0.8–1.2 mm wide.
Illustration citations
Fig. 6 View Fig , 7 View Fig ; Hooker (1844, pl. 38), reproduced here as Fig. 7 View Fig ; Dusén (1900, p. 134), as M. albiflora ; Moore (1961, p. 808), as M. pygmaea var. drucei ; Mark and Adams (1973, p. 87), as M. pygmaea s.l.; Wilson (1994, p. 245), as M. pygmaea var. drucei ; Wilson (1996, p. 221), as M. pygmaea var. drucei ; Webb and Simpson (2001, p. 142), as M. pygmaea var. drucei ; Mark (2012, p. 245), as Myosotis drucei .
Distribution
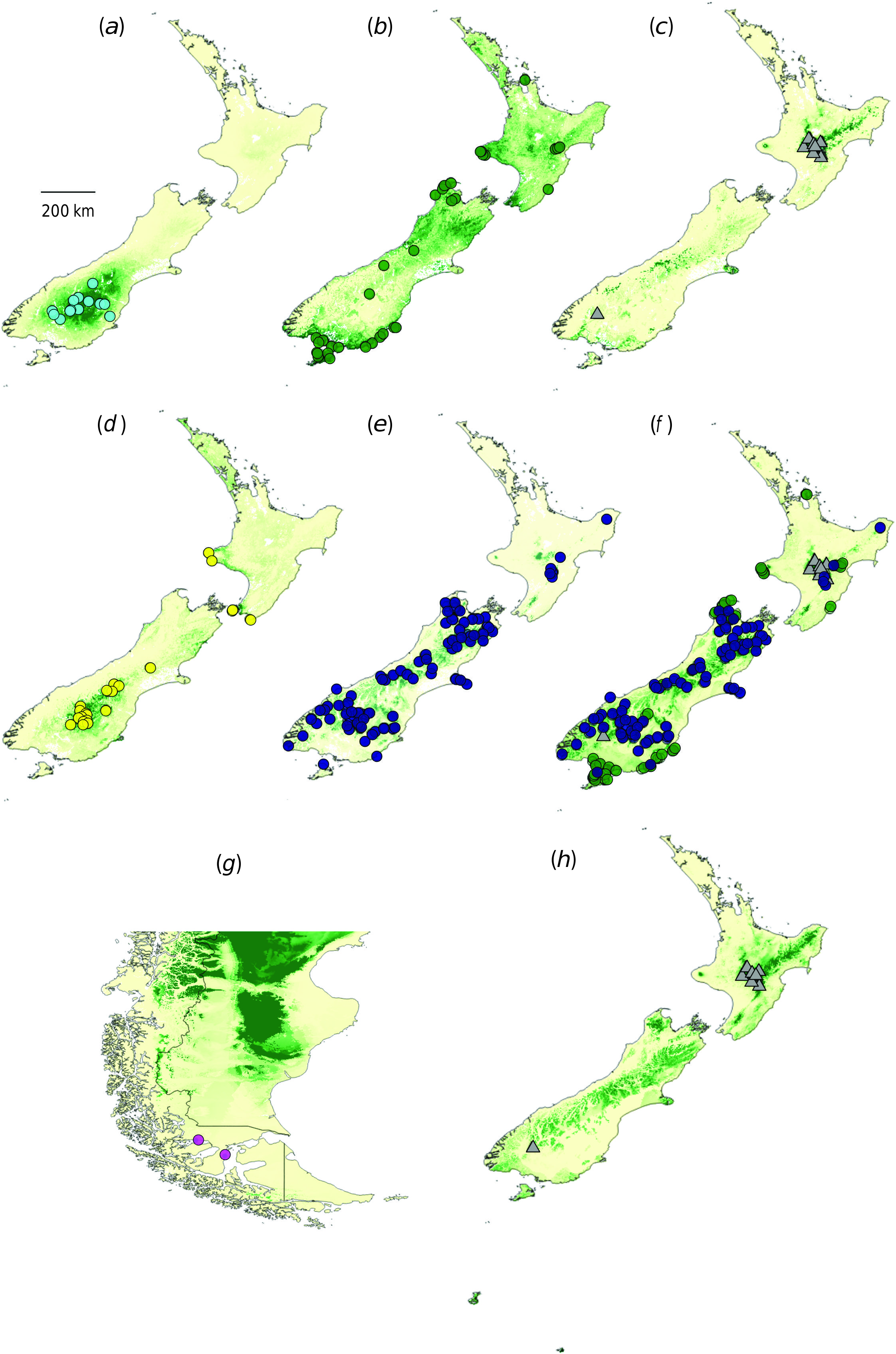
NEW ZEALAND: North Island: Gisborne, “Volcanic Plateau”, Southern North Island; South Island : Western Nelson, Sounds-Nelson, Marlborough, Westland, Canterbury, Otago, Southland, Fiordland; Stewart Island ; Campbell Island ; CHILE: Magallanes; Fig. 6 View Fig .
Habitats
From coastal turf to subalpine damp semi-stable scree, cliff faces, incised runnels and fell-fields. Elevation from sea level to 2200 m.
Phenology
Flowering August–April. Fruiting September–April. Peak flowering and fruiting December–January.
Notes
Identification. Myosotis antarctica subsp. antarctica can be distinguished from M. glauca and M. antarctica subsp. traillii on the basis of its flexuous, patent to erect trichomes. It can be separated from M. brevis because of its generally larger size, for example, corolla diameter of (1.0–)1.5–4.0 mm, calyx length at flowering of (1.2–) 2.0–3.0(–3.5) mm long and nutlets of (1.0–) 1.2–1.9 mm long (0.7–) 0.8–1.2 mm wide.
Taxonomic history. The name Myosotis antarctica was first published by Hooker (1844), for plants he collected on Campbell Island. Soon after, the name was also applied to specimens collected from elsewhere in New Zealand, for example, T. Kirk, 1877, Otago, Dart Valley (WELT SP043359) and J. Hector, s. dat. (but likely 1860s), Mount Aspiring Range (UPS V-702353). The name was first applied to Chilean specimens by Skottsberg (1915). The name M. pygmaea was published by Colenso (1884), on the basis of specimens he collected in the North Island. Although a few specimens collected from mainland New Zealand after this time were still identified as M. antarctica (e.g. Western Nelson, Mount Arthur, F. Gibbs, 1894, WELT SP002665), it became common usage to reserve the name M. antarctica for plants from Campbell Island and use M. pygmaea for all plants from the North, South and Stewart Islands, a convention which was formalised in the treatment by Moore (1961). Several varietal names within M. pygmaea were also published in the 1940s and 1960s and elevated to species rank by de Lange and Barkla in de Lange et al. (2010), i.e. M. pygmaea var. glauca was elevated to M. glauca , M. pygmaea var. minutiflora to M. brevis and M. pygmaea var. drucei to M. drucei . Two of those taxa are accepted here at species rank (see M. glauca and M. brevis above), but M. pygmaea , M. drucei and M. antarctica are considered to comprise a single species, for which M. antarctica is the earliest validly published name. The specimens that were previously identified as M. antarctica and M. drucei are considered best recognised as a single taxon at the rank of subspecies, as M. antarctica subsp. antarctica , given that they are united by morphological ( Prebble et al. 2018) but not genetic data ( Prebble et al. 2019), and they are mostly allopatric with respect to M. antarctica subsp. traillii (which encompasses specimens that were previously identified as M. pygmaea , except for the type; see below).
Not only does this circumscription of Myosotis antarctica subsp. antarctica include several previously described species, but it also subsumes two unnamed putative taxa. Those examined that are here considered to be part of this enlarged concept of M. antarctica subsp. antarctica are M. “Volcanic Plateau” (e.g. Volcanic Plateau; Kaimanawa Mountains: CHR 244442) and M. “intermedia” (e.g. Otago; Rock and Pillar Range, WELT SP089911; see table 1 in Prebble et al. 2019 for more details on these putative taxa). An additional species name, which has not been applied since its publication, M. ramificata ( Simpson 1952) , could not be distinguished morphologically from M. antarctica subsp. antarctica (fig. 6 in Prebble et al. 2018).
Patterns in the data. Specimens of Myosotis antarctica subsp. antarctica are united morphologically ( Prebble et al. 2018) but not genetically ( Prebble et al. 2019). In the nMDS analyses of morphological characters measured on herbarium specimens, all samples of M. antarctica subsp. antarctica group together (identified as M. antarctica , M. drucei , M. “intermedia” and M. “Volcanic Plateau”; fig. 6 in Prebble et al. 2018). Qualitative morphological characters found in both the herbarium and growth-room datasets distinguish M. antarctica subsp. antarctica from M. glauca and M. antarctica subsp. traillii , i.e. trichomes that are flexuous and patent to erect on the leaf blade and leaf margins. Despite the similarity in trichome types, multiple quantitative morphological characters distinguish this taxon from M. brevis (see Notes under that species).
Trichome density was the sole morphological character found to distinguish specimens identified as Myosotis drucei from an earlier narrow circumscription of M. antarctica that referred to plants from Campbell Island and Chile only. However, the ranges in trichome density overlap considerably, and so this character is not considered useful in this context ( Prebble et al. 2018). Furthermore, no morphological characters were found to distinguish specimens identified as M. “Volcanic Plateau” or M. “intermedia” from M. antarctica subsp. antarctica .
Regarding the microsatellite data, some (but not all) populations of Myosotis antarctica subsp. antarctica group together in the Structure (fig. 3 in Prebble et al. 2019) and NeighbourNet network (fig. 5 in Prebble et al. 2019) analyses. There is geographic structure in the genetic data, whereby populations growing closer together are often more closely related, but this pattern is not universal. For example, some of the populations from Central Otago group together in the Structure and NeighbourNet network (e.g. WELT SP091599, WELT SP093286 and WELT SP093291), but the populations collected from Campbell Island (WELT SP102775, WELT SP102777, WELT SP102779, and WELT SP102780) are not more closely related to each other than they are to other populations on the North and South Islands. Some of the populations identified as M. “Volcanic Plateau” do cluster together genetically (WELT SP089738, WELT SP089909; see fig. 3 in Prebble et al. 2019, K = 24), but given the lack of morphological differentiation, and the presence of geographic clustering in the genetic data discussed already, this is not considered sufficient evidence to recognise this taxon. The two populations identified as M. “intermedia” (WELT SP093292 and WELT SP100498) included in the microsatellite dataset do not cluster together genetically (fig. 3 in Prebble et al. 2019).
Pollen morphology. Pollen of Myosotis antarctica subsp. antarctica has the M. australis morphology type, the most common pollen type for bracteate-prostrate Myosotis species ( Meudt 2016) and for the ebracteate-erect species sampled so far ( Meudt et al. 2020). Representative specimens were recovered in both Clusters 1 and 2 in an nMDS analyses (see fig. 2 in Meudt 2016) along with all other specimens with pollen of the M. australis morphology type. The majority of M. antarctica subsp. antarctica pollen samples were in Cluster 1 (8 of 10), with only two (as M. sp. “intermedia”) falling into Cluster 2, one with high uncertainty. The main morphological difference between the two clusters is that those specimens in Cluster 1 usually had eight pollen apertures, and those in Cluster 2 usually had 10 apertures; however, this is not always a simple character to assign because some individuals are polymorphic for aperture number.
Chromosome number. n = 24 (CHR 101449, as Myosotis pygmaea s.l.; Beuzenberg and Hair 1983); n = 22 (AK 331000, as M. aff. drucei –M. “Volcanic Plateau”, Murray and de Lange 2013). Although this could potentially be a character used to distinguish M. “Volcanic Plateau”, we consider that we do not know enough about chromosome number variation in Myosotis to know whether this difference is meaningful, particularly given M. antarctica subsp. traillii also has a count of n = 22 (see below) and disploidy appears to be a common feature of New Zealand Myosotis on the basis of the counts undertaken so far (e.g. 2 n = 36, 40, 44, 46 and 48; Murray and de Lange 2013).
Recommended conservation status
Myosotis antarctica subsp. antarctica as circumscribed here was previously listed as two different species in de Lange et al. (2018). In that publication, it was assessed as Not Threatened (as M. drucei ) and as At Risk – Naturally Uncommon with qualifiers Data Poor, Sparse and Threatened Overseas (as M. antarctica ). Of all the pygmy forget-me-nots, M. antarctica subsp. antarctica has the highest number of estimated number of populations (299), but the smallest estimated average population size (40) ( Table 6 View Table 6 ). Taking into account evidence of census size and area of occupation, it is recommended that Myosotis antarctica subsp. antarctica maintain the same conservation status as M. antarctica in de Lange et al. (2018) , i.e. At Risk – Naturally Uncommon, Threatened Overseas (see Table 6 View Table 6 for more details).
When considering the Chilean populations and using the IUCN criteria ( IUCN Species Survival Commission 2001), Data Deficient would be the most appropriate IUCN category for Myosotis antarctica subsp. antarctica in Chile. In total, nine herbarium specimens are known from southern Chile (Magallanes Region), and these have been collected only from two locations. Six of these specimens represent two collection events of M. antarctica from Punta Arenas (Lechler s.n. 1852, S15-37467, S15-37492 and K000573650; Dusén s.n. 1895, UPS V-702363, UPS V-702365 and UPS V-702371). Therefore, the most recent collection of M. antarctica from Punta Arenas was by Per Dusén in 1895. Punta Arenas is now a city with over 100 000 inhabitants and this population and any suitable habitat may no longer exist. There is a later collection possibly from the same area but with the less precise locality information of ‘Magellans Land’ collected by Andersson in 1905 (S15-37494). By contrast, when Carl Skottsberg visited the second known location, Puerto Altamirano, and collected two specimens in 1908 (UPS V-702372 and S15-37481), he encountered ‘…a resident (at that time the only one) in Puerto Altamirano…’ ( Skottsberg 1941, p. 20), and the area is still sparsely populated today. The modelled niche for this species, projected into southern Chile, does not fit the known distribution in that region well at all, but, nevertheless, does suggest there may be additional suitable habitat for this species ( Fig. 2 View Fig ). Because the most recent herbarium specimen was collected over 100 years ago in Magallanes, M. antarctica is potentially at a severe risk of extinction there. Botanists, landowners and conservation staff working in the area are encouraged to look for populations of M. antarctica at Puerto Altamirano and other locations nearby. Further survey work is essential to determine whether the species is still extant in Chile; its population sizes, extent and quality of habitat, and population-size trends are all unknown.
Threats. Over 70% of the populations of Myosotis antarctica subsp. antarctica on the North, South and Campbell Islands are growing on DOC-managed land, and can therefore be considered protected to some degree. Additionally, some of the other populations are in reserves managed by other organisations (e.g. local councils) and therefore can also be considered protected. However, the diminutive nature of these plants, like the other pygmy forget-me-nots, means that they are at risk of weed invasion. Little is known regarding threats to the Chilean populations.
Representative specimens (415 specimens examined)
NEW ZEALAND. North Island: Gisborne: Mt Hikurangi , Jan. 1954, A. P. Druce s.n. ( CHR 86262 ). “Volcanic Plateau”: Waipakihi , 15 Feb. 2011, N. Singers s.n. ( WELT SP089738 ); Moawhango, Kaimanawa Ranges , Jan. 1974, A. P. Druce s.n. ( CHR 252337 ); Mt Tihia , South West of Lake Taupo , Apr. 1974, A. P. Druce s.n. ( CHR 273143 ). Southern North Island : Ruahine Range , Te Hekenga , Feb. 1968, A. P. Druce s.n. ( CHR 190683 ); Ruahine Range , Makirikiri tarns , Jan. 1977, A. P. Druce s.n. ( CHR 310160 ). South Island : Marlborough: Tapuae-o-Uenuku , Feb. 1981, B. Molloy s.n. ( CHR 386879 ); Mt Altimarlock , 8 Feb. 2013, J. M. Prebble JMP13026 & C. Jones ( WELT SP100428 ). Western Nelson, Cobb Valley , near Lake Sylvester , 13 Jan. 1962, R. Melville 5961 & H. Telbot ( CHR 142854 ). Canterbury: Banks Peninsula , Port Hills , Trig O, 19 Feb. 2012, J. M. Prebble JMP12007 ( WELT SP093292 ). Otago, Macraes flat , 1 Mar. 2013, J. M. Prebble JMP13040 & K. Pilkington ( WELT SP100498 ); Taieri River , Beaumont Station , 15 Dec. 2009, M. Thorsen 129/09 ( WELT SP089916 ); Rock and Pillar Range , 2 Feb. 2014, J. M. Prebble JMP14002 & E. Connor ( WELT SP102783 ). Fiordland: Beansburn , s. dat., B. D. Rance s.n. ( WELT SP104524 ). Stewart Island / Rakiura: Mt Anglem , 6 Jan. 2000, P. J. de Lange 4109, ( AK 251910 ). Campbell Island : Mt Azimuth ; 27 Dec. 2013, J. M. Prebble JMP13065 & A. J. Fergus ( WELT SP102777 ); Mt Honey , 28 Dec. 2013, J. M. Prebble JMP13067 & A. J. Fergus ( WELT SP102779 ) and JMP13068 & A. J. Fergus ( WELT SP102780 ) . CHILE: Magallanes: Punta Arenas , 12 Dec. 1895, P. K. H. Dusén 173 ( UPS V-702363 ); Skyring Sound, Puerto Altamirano, 22 Apr. 1908, C. Skottsberg s.n. ( UPS V-702372) .

| CHR |
CHR |
| WELT |
WELT |
| UPS |
UPS |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Myosotis antarctica Hook.f. subsp. antarctica
| Prebble, Jessica M., Symonds, V. Vaughan, Tate, Jennifer A. & Meudt, Heidi M. 2022 |
Myosotis ramificata G.Simpson, Trans. & Proc. Roy. Soc.
| 1952: 426 |
Myosotis pygmaea
| Colenso, Trans. & Proc. 1884: 334 |