
Potamocypris compressa Furtos, 1933
|
publication ID |
https://doi.org/10.5281/zenodo.205489 |
|
DOI |
https://doi.org/10.5281/zenodo.5681362 |
|
persistent identifier |
https://treatment.plazi.org/id/03AF0674-C366-FFC8-FF55-AEC9FD153DE4 |
|
treatment provided by |
Plazi |
|
scientific name |
Potamocypris compressa Furtos, 1933 |
| status |
|
Potamocypris compressa Furtos, 1933
( Figs 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
1933 Potamocypris smaragdina View in CoL var. compressa n. var. Furtos, 435–437, pl. 6, figs 10–14. 1941 Potamocypris smaragdina compressa Furtos, 1933 ; Dobbin, 231–232, pl. 2, figs 1–6.
Material examined. Several hundred adult and juvenile specimens collected alive from seven rock pools in the Hidden Canyon area, near Moab, Utah, USA (see Table 1 for details).
Type locality. Rock pool, Starve Island, Lake Erie, Ohio, USA; latitude 41 37ʹ 36ʺ N, longitude 82 49ʹ 21ʺ W, elevation 173 m. Furtos (1933) did not designate a type locality but this is the first of the nine sites she mentioned and also one that she singled out as being where this species was “particularly characteristic”.
Type material. Division of Crustacea, National Museum of Natural History, Smithsonian Institution, USA; USNM 6793, 100 syntypes ( Harrison 1987).
Diagnosis. A hirsute Potamocypris ; in lateral view, Cp with maximum height close to or slightly in front of mid-length; in dorsal view, RV overlapping LV anteriorly, LV overlapping RV posteriorly; valves ornamented with fine pits / punctae arranged in a faint reticulum (more clearly expressed in juveniles). Marginal zone of LV: selvage peripheral at anterior end, displaced inwards at posterior end. Long swimming setae on the A2, extending substantially beyond the tips of the terminal claws of the endopodite; Mx palp with four spines on the distal podomere; L5 branchial plate with two rays; lateral shield of hemipenis subtrapezoidal, beak-like.
Abbreviated description. Valves medium-sized (Le = c. 610 µm), distinctly asymmetrical; dimorphic, female ( Figs 1 View FIGURE 1 H, I; 4A, B) larger than male ( Figs 1 View FIGURE 1 J, K; 4C). Female LV overreaching RV posteriorly ( Fig. 1 View FIGURE 1 D, F, G). In lateral view female LV ( Figs 1 View FIGURE 1 A, H; 4B) rounded subtriangular, moderately elongate, outer margin with a short, almost straight section at the posteroventral corner where it overreaches the RV. In lateral view female RV ( Figs 1 View FIGURE 1 I; 4A) rounded subtriangular, dorsally more highly arched than LV, overreaching LV anteriorly ( Fig. 1 View FIGURE 1 D, E, G) and dorsally ( Fig. 1 View FIGURE 1 A), posteroventral corner evenly rounded. In both sexes maximum height close to or slightly in front of mid-length in LV and RV, maximum width of Cp close to mid-length. Calcified inner lamella of LV relatively narrow, selvage peripheral anteriorly and displaced inwards posteroventrally, inner list and outer marginal flange present posteroventrally ( Fig. 1 View FIGURE 1 H, J). Calcified inner lamella of RV relatively narrow, selvage peripheral along entire free margin of valve ( Fig. 1 View FIGURE 1 I, K). Male valves with the same features as those of females but smaller and with a narrower posterior marginal flange. External surface of valves with fine pitting / punctae arranged in a faint reticulate pattern and hirsute (i.e. with abundant sensilla).
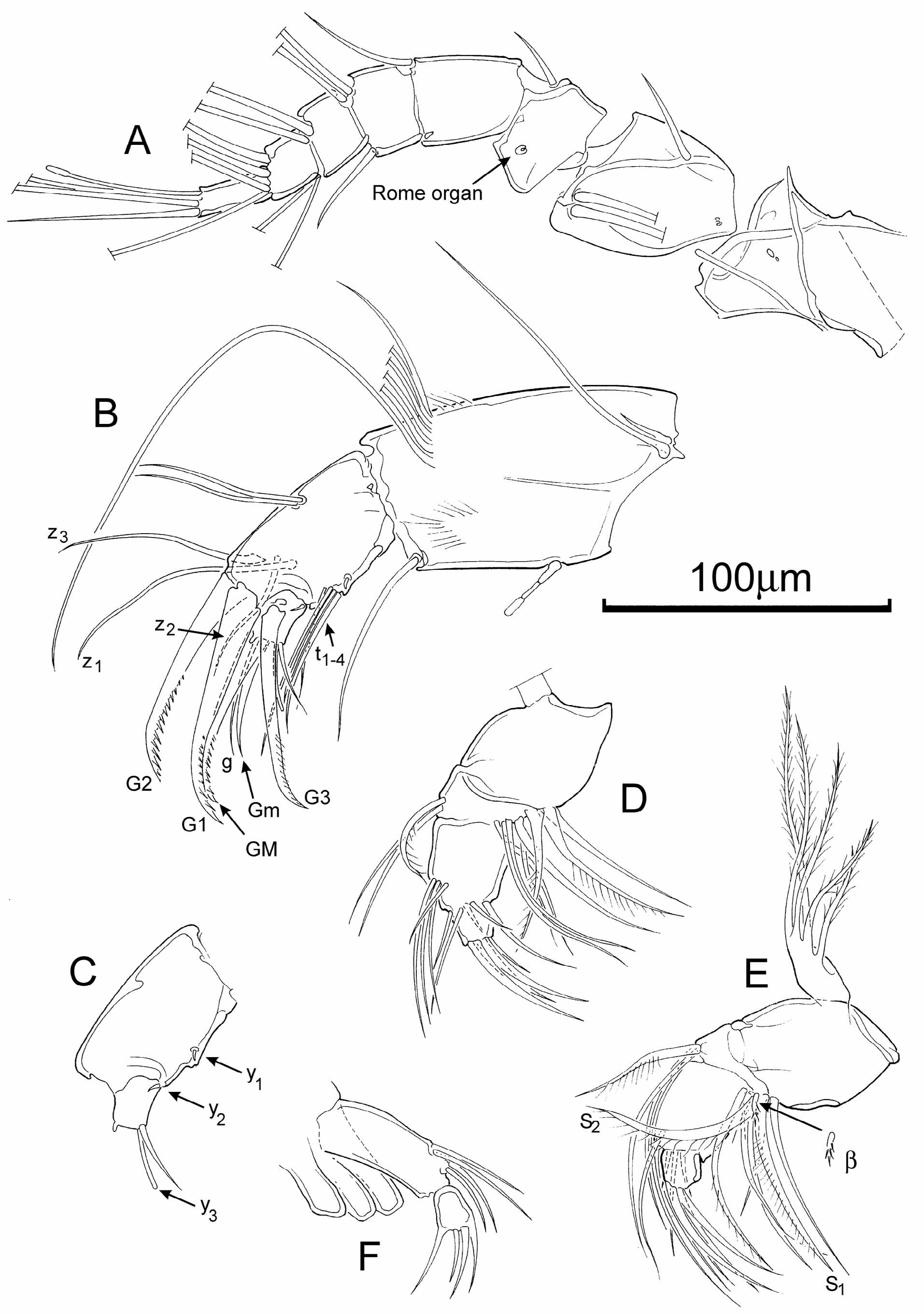
A1 ( Fig. 2 View FIGURE 2 A) with long natatory setae and seven articulated podomeres (the basal/proximal podomere possibly split into two segments). Setation of podomeres from first (proximal) to seventh (distal): first with a pair of long setae and one short seta; second with one short seta; third with one short seta; fourth with two long and one short setae; fifth and sixth each with four long setae; seventh with three long and one short setae and an aesthetasc. Small rounded Rome organ on second podomere. Wouters organ not seen, even at highest magnification.
A2 ( Fig. 2 View FIGURE 2 B, C) with long natatory setae which extend beyond the ends of the terminal claws by approximately the length of the claws (or about 30% of the length of the swimming setae). Penultimate podomere with four t-setae and three z-setae (one short and two long). Four aesthetascs: Y (large), y1 and y2 (small) and y3 (moderately large). Terminal claws all set with spines in their distal half.
Md palp ( Fig. 2 View FIGURE 2 D, E) with three terminal claws, alpha seta slender and short, beta-seta swollen and hirsute, gamma-seta slender and short, not claw-like; exopodite with five rays.
Mx palp ( Fig. 2 View FIGURE 2 F) with two podomeres, the distal one spatulate with four small terminal claws.
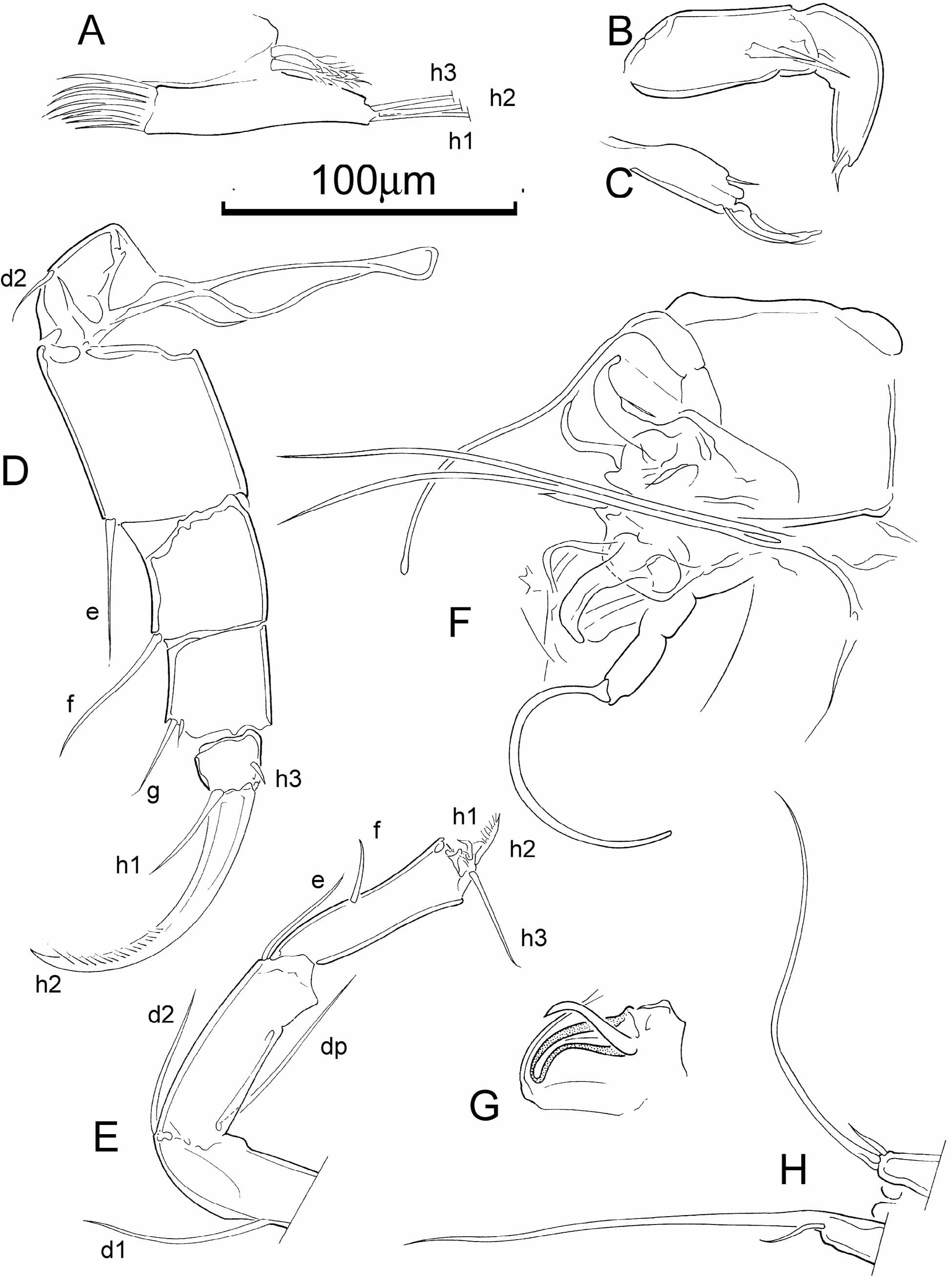
L5 dimorphic: female palps symmetrical with three terminal setae (h1–3) ( Fig. 3 View FIGURE 3 A), male palps asymmetrical claspers ( Fig. 3 View FIGURE 3 B, C); in both sexes branchial plate with two rays.
L6 (walking leg) ( Fig. 3 View FIGURE 3 D) large, robust, with five articulated podomeres; terminal podomere apically bearing a large claw (h2) set with spines on its distal half, a smaller seta (h1), and a small subapical seta (h3); penultimate podomere with two apical setae of unequal size.
L7 (cleaning limb) ( Fig. 3 View FIGURE 3 E) slender with four articulated podomeres, the terminal one very short, with one long (h3), one very short (h2) and one short claw-like (h1) setae, and forming a pincer with the terminal part of the penultimate podomere; basal podomere with three long setae, two apical (d2 and dp), one more proximal (d1).
Uropods (also known as caudal rami, and formerly as the furca) ( Fig. 3 View FIGURE 3 H) each with a short conical base bearing a short seta and a long, flagellum-like seta.
Female reproductive organs with robust genital hooks ( Fig. 3 View FIGURE 3 F, G) and large spiral ducts (the latter seen uncoiled in Fig. 3 View FIGURE 3 F).
Male hemipenis ( Fig. 4 View FIGURE 4 D, E) with cypridopsine-type labyrinth and subtrapezoidal, beak-shaped lateral shield (distal lobe); Zenker’s Organ ( Fig. 4 View FIGURE 4 F) with approximately 12 spinose whorls.
Figured specimens. OC.3154 ( Fig. 1 View FIGURE 1 A), female Cp; OC.3155 ( Fig. 1 View FIGURE 1 B), juvenile A-1 Cp; OC.3156 ( Fig. 1 View FIGURE 1 C), juvenile A-2 Cp; OC3157 ( Figs 3 View FIGURE 3 B, C), male RV and LV separated, appendages dissected; OC.3158 ( Figs 3 View FIGURE 3 D; 4A, B), female RV and LV separated, appendages dissected; OC.3159 ( Fig. 1 View FIGURE 1 D, E, F), female Cp; OC.3160 ( Fig. 1 View FIGURE 1 G), female Cp; OC.3161 ( Fig. 4 View FIGURE 4 C, F), male RV and LV separated, appendages dissected; OC.3162 ( Figs 2 View FIGURE 2 F; 3A, F, E), female RV and LV separated, appendages dissected; OC.3163 ( Figs 1 View FIGURE 1 H, I; 3G, H), female RV and LV separated, appendages dissected; OC.3164 ( Figs 1 View FIGURE 1 J, K; 4D, E), male RV and LV separated, appendages dissected; OC.3165 ( Fig. 2 View FIGURE 2 D, E), female RV and LV separated, appendages dissected. All from rock pools in the Hidden Canyon area, Utah, USA; numbers OC.3154–3160 are from pool HC3-R4, OC.3161–3165 are from pool HC2-RB (see tables 1 and 2).
Ecology. Our specimens were collected alive from a range of ephemeral to “more permanent” rock pools, representing a continuum of hydroperiods, in low conductivity fresh waters with pH>9 (see Table 2; note that the values given were measured during the day at the time of collection; values may drop as low as pH 4 at night). Furtos (1933) reported the species as being common in shallow rock pools and in weedy inlets of Lake Erie, as well as in smaller lakes; in a pool on Starve Island, Lake Erie (designated herein as the type locality) she found it thriving from mid-May to November in a “practically pure culture”.
Distribution. North America, so far confirmed only from the three areas reported herein ( Fig. 5 View FIGURE 5 ); unknown in Europe. Furtos (1933) recorded it from the environs of Lake Erie (in pools on islands and on the coast) and in four other lakes in Ohio.
Discussion. Potamocypris smaragdina ( Vávra, 1891) (as redescribed and illustrated by Meisch 1985; see also Meisch 2000) is similar to our specimens but has a less elongate carapace, with maximum height distinctly anterior of mid-length, and in dorsal view the LV overlaps the RV at both anterior and posterior ends. In the LV of our specimens the anterior selvage is peripheral, but in P. smaragdina it is displaced inwards, exposing a well-developed anterior marginal flange. Our specimens match those described and illustrated as P. smaragdina var. compressa by Furtos (1933), who did not mention males. This variety was also noted by Dobbin (1941) as a subspecies, from Little San Poil Lake in Ferry County, Washington; she stated that males were unknown. Hoff (1942) considered that Furtos (1933), in describing the new variety compressa , had been misled by errors in Vávra’s original description of P. smaragdina from Europe, and concluded that there were no significant differences between the European species and Furtos’ variety from Ohio. However, Hoff only considered the carapace dimensions (length, height, width) and the length of the A2 swimming setae relative to the terminal claws, and paid no attention to the shape of the carapace (maximum height well in front of mid-length in P. smaragdina , close to mid-length in P. smaragdina var. compressa ), or the overlap relationships of the valves. The male of P. smaragdina as illustrated by Hoff (1942) has a hemipenis with a lateral shield that is bilobate with rounded lobes, while that of our P. compressa is more trapezoidal, expanded proximally, and distally with rounded corners, one an obtuse angle and one acute. Clearly our material is not conspecific with P. smaragdina as described and illustrated by Vávra (1891) and Hoff (1942). Furtos’ description and illustrations indicated that the maximum height of the carapace is somewhat further in front of mid-length than in our material, but carapace shape in lateral view is known to be highly variable in other Potamocypris species (see, e.g., Meisch 1985) and we do not consider this character to be as significant as valve overlap relationships or soft-part morphology. Otherwise, our specimens match Furtos' description and illustrations in every respect except colour: she described hers as yellowish-green with two darker green dorso-lateral stripes (which corresponds to the description of P. smaragdina ; see, e.g., Meisch, 2000), whereas ours seem to a more uniform medium green. Colour patterns may vary within species, however, and it must be borne in mind that although they are still distinctly coloured, some colour change may have occurred in our specimens since collection, due to preservation in ethanol. Moreover, Furtos commented that the strength of colour of the stripes varied from very faint to brilliant. Dobbin (1941) reported finding P. smaragdina var. compressa in Washington. Meisch (1985) commented that if Dobbin’s indication of two branchial plate rays on the L5 is correct, her specimens must be a different species (i.e., not P. smaragdina which has only one L5 ray); Dobbin illustrated the distal podomere of the Mx endopodite as having four spines, the same as in our specimens, which likewise have two L5 rays, and we consider Dobbin’s specimens to belong to P. compressa .
Potamocypris pallida Alm, 1914 has a similar carapace shape to P. compressa , with RV overlapping LV anteriorly and LV overlapping RV posteriorly, but is not so hirsute and has short swimming setae on the A2.
Potamocypris islagrandensis Hoff, 1943 , from Louisiana, has a similar lateral outline to P. compressa but in dorsal view has the opposite valve overlap relationship and is widest behind mid-length; moreover, it has only one L5 branchial plate ray and the lateral shield of the hemipenis is broadly bilobate and rounded ( Hoff 1943).
European P. unicaudata Schäfer, 1943 View in CoL , as illustrated by Meisch (1985; 2000) (who says that males are unknown), is also similar to P. compressa but has maximum carapace height distinctly in front of mid-length and a rather more pronounced posteroventral valve overlap so that in dorsal view the valves are more distinctly asymmetrical. Furthermore, like P. smaragdina View in CoL , P. unicaudata View in CoL has only one branchial plate ray on the L5 and four spines on the distal podomere of the Mx endopodite ( P. compressa also has four spines, but has two L5 rays).
Potamocypris villosa (Jurine, 1820) is another similar, hirsute species, with two L5 branchial plate rays, but (according to the description and illustrations of Meisch 1985) is relatively less elongate, has five spines on the terminal podomere of the Mx endopodite, and the A2 swimming setae tend to be relatively shorter, only overlapping the A2 claws by <20% of the setal length (although this is somewhat variable). Furthermore, males of P. villosa were described from Europe by Martens & Meisch (1985), who illustrated a male hemipenis with a quite differently-shaped lateral shield expanding distally, with a broadly rounded lobe and a small beak-like lobe.
Potamocypris arcuata (Sars, 1903) is also of similar shape and hirsute, and has distinct pits on the valves; these pits (as illustrated by Meisch 1985) are relatively larger and more spaced out (inter-pit space typically two or three times the pit diameter) than those on our specimens of P. compressa , which are very fine and closely spaced, forming polygonal patterns. Such apparent differences in surface ornament may be of limited value as species-diagnostic characters, however, they may represent ecophenotypic variation within species. The valve overlap relations in dorsal view are also similar although it must be noted that, according to Meisch (1985), the anterior overlap in P. arcuata is rather variable. The shape in lateral view of P. arcuata is also quite variable, but generally it seems less elongate than P. compressa . In dorsal view P. arcuata is more compressed than P. compressa . Potamocypris arcuata has five spines on the distal podomere of the Mx endopodite, whereas P. compressa has four. Males of P. a rc u - ata have been reported from North Africa by Gauthier (1928) who illustrated a hemipenis with a relatively short, distally expanded lateral shield, with one broadly rounded and one acutely rounded distal corner; it does not match the hemipenis shape of P. compressa .
We agree with Meisch (1985) that P. c o m o s a, described by Furtos (1933) from the shores of Lake Erie, may be synonymous with P. smaragdina View in CoL . It certainly matches well, including having the LV extending beyond the RV anteriorly and posteriorly in dorsal view. The hemipenis, described by Furtos as "quadrangular, the lateral lobe expanded distally" (op. cit., p. 435) is illustrated (op. cit., fig. 20) as having a bilobate lateral shield that corresponds closely to that illustrated by Hoff (1942) for the male of P. smaragdina View in CoL . No males of P. smaragdina View in CoL have been found in Europe, and Meisch (1985) does not discuss the morphology of males, merely commenting that males have been reported from North America by Hoff (1942) and Ferguson (1944).
Delorme’s (1970) identification of P. smaragdina ( Vávra, 1891) View in CoL in Canada is probably correct; the form he illustrates is hirsute and appears to have similar carapace features to European P. smaragdina View in CoL , including the left valve overlapping the right both anteriorly and posteriorly, and the maximum height being in front of mid-length, although the colour is simply stated to be green with no mention of banding. However, although the specimens figured by Delorme (1970) can be assigned with confidence to P. smaragdina View in CoL , it remains possible that his material may have included populations that we would now assign to P. compressa .
Meisch (1985) noted that the record of P. smaragdina View in CoL (including males) from Argentina by Ramirez (1967) probably belongs to a different species. We agree with Meisch's opinion; the species illustrated by Ramirez appears to have a more elongate carapace, has two L5 branchial plate rays, five spines on the distal podomere of the Mx endopodite, and the lateral shield of the male hemipenis is distally expanded into two angular lobes, each with an acute angular outer corner, the last two features also serving to distinguish it from P. c o m p re s s a. We consider it likely that Ramirez’ material represents a new species.
We have been unable to determine any fossil record of P. smaragdina View in CoL . Swain (1999) included P. smaragdina View in CoL var. compressa in the synonymy of P. smaragdina View in CoL , for which he noted a number of Pleistocene records in the USA. As with the living records, therefore, it remains possible that fossil records of P. compressa are hidden because it has not been recognised hitherto as a valid species.



| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Cypridopsinae |
|
Genus |
Potamocypris compressa Furtos, 1933
| Horne, David J., Jocque, Merlijn, Brendonck, Luc & Martens, Koen 2011 |
Potamocypris islagrandensis
| Hoff 1943 |
P. unicaudata Schäfer, 1943
| Schafer 1943 |
Potamocypris smaragdina compressa
| Furtos 1933 |
Potamocypris arcuata
| Sars 1903 |
P. smaragdina ( Vávra, 1891 )
| Vavra 1891 |
Potamocypris villosa
| Jurine 1820 |