
Feuerborniella veracruzana IbáñezBernal, 2004
|
publication ID |
https://doi.org/10.11646/zootaxa.412.1.1 |
|
DOI |
https://doi.org/10.5281/zenodo.5228236 |
|
persistent identifier |
https://treatment.plazi.org/id/03AF1D19-FFDF-FF99-9B04-822DFB7E6526 |
|
treatment provided by |
Felipe |
|
scientific name |
Feuerborniella veracruzana IbáñezBernal |
| status |
sp. nov. |
Feuerborniella veracruzana IbáñezBernal View in CoL , sp. nov.
( Figs. 1–15 View FIGURE View FIGURE 8–15 )
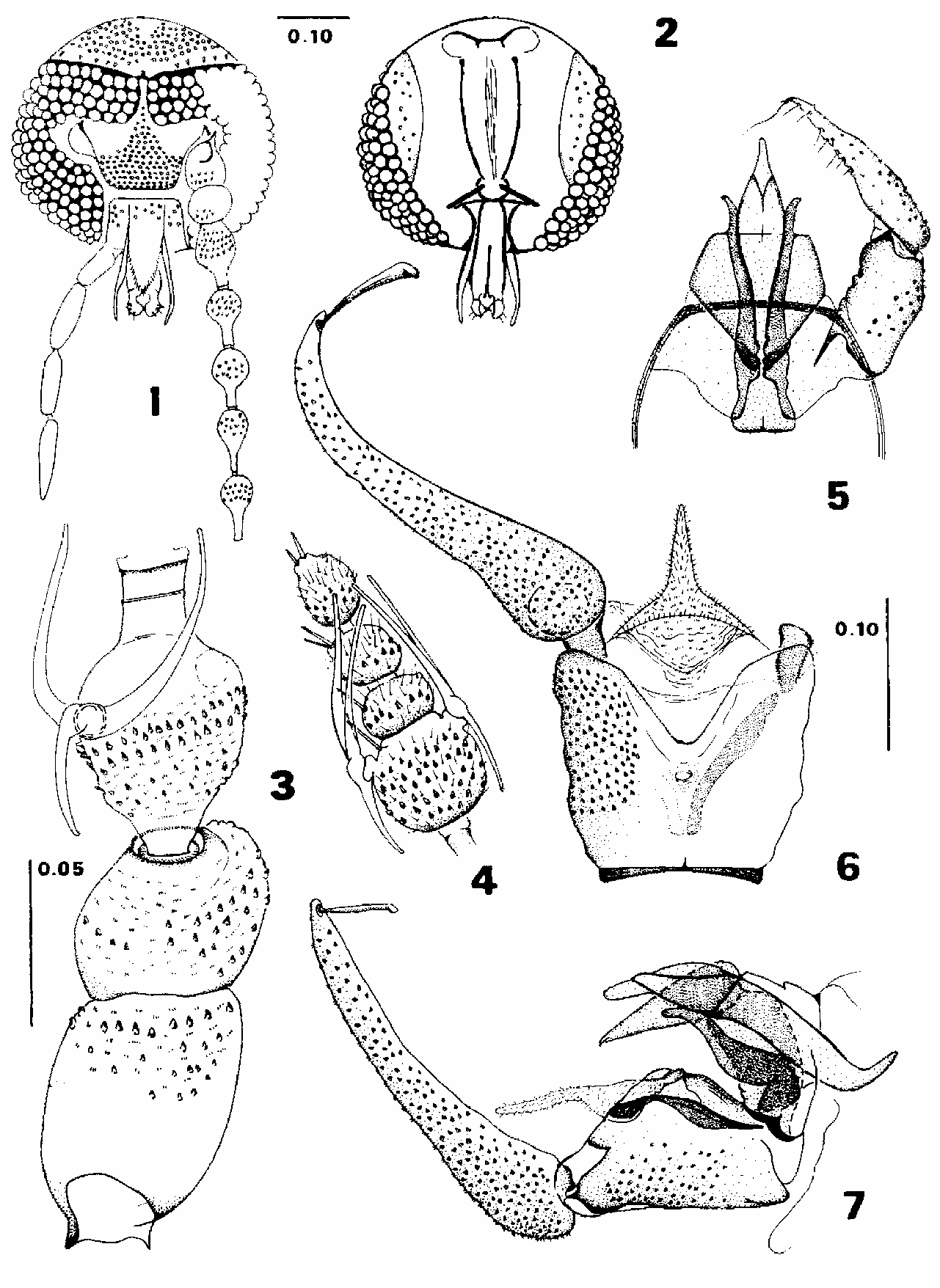
Male description ( Figs. 1–7 View FIGURE ). Body vestiture apparently uniform pale gray, the integument yellowish brown. Head ( Fig. 1–2 View FIGURE ): Eyes separated by one or one and a half facet diameters; eyebridge with four rows of facets; interocular suture somewhat arched and with a poorly visible dorsal middlemost spur; a row of five setae along the upper margin of each eye; the vertex rounded in outline, with a single median patch of scale alveoli leaving a small naked area in the upper central area which extends downward without reaching the level of the interocular suture; frontal patch of setae alveoli undivided, extending upward between the eyes to reach the level of the third facet ( Fig. 1 View FIGURE ). Ascendant tentorial arms straight, narrow, as long as 8x its width; posterior tentorial arms composed of a pair of laterally sclerotized, nearly parallel, concave flanges in posterior view; maxillary apodeme nearly straight; posterior margin of the eyes sigmoidal in outline, and a postoccipital plate well separated by a suture ( Fig. 2 View FIGURE ). Labellum bulbous with three or four long anteapical setae and two to four short spiniform setae in the internal margin near the apex, but without blunt teeth (as in Fig. 9 View FIGURE 8–15 ). Palpus as long as 0.36x antenna length, and about 1.1x as long as head height; proportion of palpomeres: 1.00: 1.42: 1.36: 1.46 ( Fig. 1 View FIGURE ). Antennae as long as 0.5x wing length, with 14 flagellomeres; scape barrelshaped, longer than the spherical pedicel; flagellomere 1 with node 2x as long as its own internode; subsequent flagellomeres similar to 1, but nodes progressively decreasing in size and internodes increasing in length to such an extent that flagellomere 10 has the node equal in length to the internode; each internode parallel sided, uniformly sclerotized but with a median transverse sulcus ( Fig. 3 View FIGURE ); flagellomere 11 without internode; flagellomeres 12–14 small and globular, not fused, 12–13 nearly sessile but in a short broad pedunculus, 14 with a long pedunculus; a pair of “Y”shaped ascoids present on flagellomeres 1–11, each with two large anterior branches and one shorter posterior branch; the last three flagellomeres only with the short spiniform sensilla ( Fig. 4 View FIGURE ). Thorax: Wing length about 2.5x breadth; Sc ending at the level of the base of R 1; R 5 ending at wing tip; base of M 2 not connected; wing membrane without hairs; proportion of R 2+3:R 3 = 2.9:2.8; M 1+2:M 2 = 1.5:4.0. Genitalia with elaborate and symmetric aedeagus ( Figs. 5–7 View FIGURE ). Cercopod with setae alveoli covering its surface, as long as 1.7x the lateral length of epandrium, bearing a clavate retinaculum which is as long as 0.3x the cercopod length; epandrium strongly bilobed apically, a little longer than wide, with two well separated lateral patches that do not reach the proximal portion and with a small pseudospiracular opening ( Fig. 6 View FIGURE ). Gonocoxite subcylindrical, with an external lateral patch of setae alveoli, 0.64x as long as the gonostylus; gonocoxal apodemes broad and platelike meeting beneath the aedeagal apodeme, only slightly expanded, with the internal margin nearly straight and convergent posteriorly. Gonostylus hirsute, straight, but ending in a sharp, entally curved apex, and with an external basal patch of setae alveoli and an irregular row of small setae in the internal apical margin ( Fig. View FIGURE 5). Aedeagus fusiform, with the apex acutely rounded in dorsoventral view ( Fig. 5 View FIGURE ), canoelike in profile ( Fig. 7 View FIGURE ), flanked by one pair of relatively long and nearly straight lateral shafts, except for a slightly ectally directed curvature of the apical portion; apex of lateral shafts reaching the apical fourth of aedeagus ( Fig. 5 View FIGURE ); aedeagal apodeme broad and truncated; aedeagal sheath trapezoidal in dorsoventral view. The hypandrium is slender and diademlike ( Fig. 5 View FIGURE ). Tergite 10 is globular with a long conical median fleshy projection ( Figs. 6–7 View FIGURE ).
Measurements. Head height: 0.315 (0.31–0.32) n=2; proboscis length: 0.08; palpus length: 0.35 (0.35–0.37) n=4; antenna length: 0.98 (0.94–1.03) n=5; wing length: 1.91 (1.71–2.25) n=5; wing width: 0.77 (0.68–0.76) n=5; cercopod length: 0.28 (0.27–0.30) n=5; gonostylus length: 0.13 (0.13–0.14) n=5; gonocoxite length: 0.08 (0.08–0.09) n=5; aedeagus length (from apodeme): 0.19 (0.17–0.21) n=5.
Female description ( Figs. 8–15 View FIGURE 8–15 ). Similar to the male except for the following features: palpus 0.36x as long as antenna; proportion of palpomeres: 1.0:1.2:1.2:1.3.
Antennae ( Figs. 11–12 View FIGURE 8–15 ) 0.46x as long as wing; flagellomeres 12–14 each with a longer petiole ( Fig. 12 View FIGURE 8–15 ). Wing 2.7x as long as width; proportion of R2+3:R3=1.0:1.0; M1+2:M2=1.0:2.2 ( Fig. 10 View FIGURE 8–15 ). Genitalia ( Figs. 13–15 View FIGURE 8–15 ): subgenital plate 0.67x as long as broad; distal lobes “X”shaped, each with one sensory setae originating near the basal margin of the lobes at the level of the triangular genital sac ( Fig. 14 View FIGURE 8–15 ). Spermathecae complex as figured ( Fig. 13 View FIGURE 8–15 ); corpus spermathecae sculptured; median arms bladelike; posterior arms slightly divergent posteriorly; cercus long and gradually tapered ( Fig. 15 View FIGURE 8–15 ).
Measurements. Head height: 0.315 (0.31–0.32) n=2; proboscis length: 0.08, n=2; palpus length: 0.36 (0.35–0.37) n=2; antenna length: 1.0; wing length: 1.99 (1.89–2.09) n=2; wing width: 0745 (0.73–0.76) n=2; cercus length: 0.24 (0.21–0.27) n=2.
Immature stages: unknown.
Etymology: veracruzana , pertaining to Veracruz State, Mexico.
Type locality. Mexico, Veracruz, Tlilapan, Cueva Matzinga. The specimens were obtained by aspirating resting adults from the cave walls, somewhat beyond the cave entrance.
Holotype male, allotype female, and three males and one female paratypes (as labeled): Mexico, Veracruz, Tlilapan, Cueva Matzinga , 25x1989, F. Lledías, col. All specimens are mounted on slides with Canada balsam . Holotype, allotype and two male paratypes are deposited in the entomological collection at the Instituto de Ecología , A.C. ( IEXA), Xalapa, Veracruz, Mexico . One male and one female paratype are deposited in the Los Angeles County Museum of Natural History ( LACMNH), California, USA .
Comments. Eaton (1904) erected the subgenus Philosepedon within Psychoda Latreille , based on the presence of two retinacula in the male cercopodium. Quate (1959) maintained that this taxon is a valid genus and gave the diagnostic characteristics for its recognition based on the presence of a bulbous labellum without teeth, 16 antennomeres (scape, pedicel and 14 flagellomeres) with the apical three reduced, ascoids “Y”shaped, wing with radial fork distad of the median fork by a distance equal to more than 3x the width of cell R 2 at the point of bifurcation, male with cercopodium not much longer than epandrium, with 2 or 3 retinacula (some species with only one), aedeagus symmetric, with one or two pairs of lateral shafts, female genitalia without digit and cercus short, equal or shorter than the subgenital plate.
Botosaneanu and Vaillant (1970) erected the genus Quatiella based on the discovery of the species Q. cubana Botosaneanu and Vaillant. They used the following generic characteristics: eyes approximate but not contiguous, bulbous labellum with setae and short spines (without blunt teeth), 16 antennomeres with the terminal three reduced, ascoids trifid, male genitalia with cercopodium bearing one retinaculum, aedeagus symmetric, female genitalia with a very short cercus. Additionally, they denoted in a short complementary key for that refered by them as published in 1970, actually published one year later ( Vaillant, 1971), that epadrium is much longer than wide and that the base of the gonopods are contiguous with each other, characteristics used to separate Quatiella from Feuerborniella . Based on the figure of the wing presented by Botosaneanu and Vaillant (1970, p. 175) and the figure of the typespecies Philosepedon leonhardti Vaillant (= Quatiella interdicta Dyar ) ( Vaillant 1968), we can infer that at least these species have the radial fork distad of the median fork by a distance about the width of cell R 2 at the point of bifurcation.
Vaillant (1973) later described Philosepedon quatei and presented a key to the North American genera of his Threticus group. In that work, the distinctive characteristics between Philosepedon and Quatiella were: cercopodium with 2 or 2 groups of retinacula versus only one retinaculum, respectively. In the same year, Duckhouse (1973) placed Quatiella and Eurygarka Quate as subgenera of Philosepedon , considering P. interdicta under the subgenus Philosepedon . In addition, the Brazilian species he originally described as Psychoda plaumanni , Psychoda spathipenis ( Duckhouse, 1968) , and Psychoda fumata Knab (known from Mexico) were also included. In another publication, Duckhouse (1974) expressed his opinion about the validity of Quatiella and concluded that there was no justification to retain it as a separate genus. The only conspicuous difference between Eurygarka and the others, as was first noted by Quate (1959) is the form of the antennal ascoids, whereas the other diagnostic characteristics overlap or are not sufficiently precise to separate them.
Vaillant (1974) described the genus Feuerborniella to harbor the Palearctic species F. obscura (Tonnoir) , and considered both Brazilian species of Duckhouse and Trichopsychoda malayensis Satchell , under this genus. Vaillant considered Threticus Eaton , Philosepedon , Quatiella , Nielseniella Vaillant , Feuerborniella , and three no longer valid genera as the Threticus group of Telmatoscopini ( Vaillant 1973, 1974). It is important to note that the original description of Feuerborniella was published 4 years after the first mention of the genus by Botosaneanu and Vaillant (1970), situation that could invalidate the name in accordance with ICNZ (2000).
Vaillant (1990) retained Threticus , Philosepedon , Trichopsychoda Tonnoir , Feuerborniella , Quatiella and Nielseniella as valid genera under the tribe Mormiini Enderlein , corresponding to the tribe Telmatoscopini of Vaillant (1971), or to the tribes Paramormiini and part of Psychodini of Duckhouse (1987). More recently, Wagner (1991) considered Feuerborniella , Nielseniella , Trichopsychoda , Threticus and Philosepedon as part of the tribe Telmatoscopini .
Duckhouse (1985) redefined the tribe Psychodini based on the following apomorphies: antennal flagellomeres flaskshaped except the terminal three which are reduced and without ascoids; ascoids Yshaped or derived from this type by loss of one or two anterior branches; wing pointed with R 5 ending at the apex; subcosta (Sc) short and thick; wings folding in tectiform position; erect hair present on R 1, and epandrium of male with single pseudospiracular opening, as well as other characters observed in larvae. The simple characteristic of the flattened labellum is insufficient to separate the tribe, because the species of Threticus have both conditions of the labellum. Duckhouse (1985) included the genera Psychoda , Feuerborniella , Neomaruina Vaillant , Epacreton Quate , Philosepedon , Trichopsychoda and Threticus in tribe Psychodini . Other genera that have been included more recently in the Psychodini are: Chodopsycha Jezek , Logima Eaton , Psycha Jezek , Psychodocha Jezek , and Psychomora Jezek ( Duckhouse and Lewis, 2000) . These inclusions are well supported based on the available information and we follows the concept provided by Duckhouse (1985) on the tribe Psychodini .
The new species described in this report correspond to the genus Feuerborniella by the approximate eyes, ocular bridge with 4 rows of facets, 14 flagellomeres, “Y”shaped ascoids, apical 3 flagellomeres reduced in size, labellum small and fleshy with setae and small spines but without blunt teeth, R 5 ending at the wing apex, males with cercopod longer than epandrium, bearing only one simple (but not considerably long) retinaculum, gonocoxites separated from each other, and female genitalia with long, gradually tapered cercus and without digit ( Vaillant 1974). It is important to note that Philosepedon spp. may have two or two groups of cercopodial retinacula in the male and the cercus short in the female. Eurygarka spp. are characterized by the foliar sulcate branches of the “V”shaped antennal ascoids. Quatiella spp. have the gonocoxites contiguous and epandrium considerably longer than wide in the male, and short cerci in the female. Threticus spp. have 3 or 3 groups of cercopodial retinacula and the aedeagus asymmetric. Trichopsychoda spp. are characterized by the long, thin retinacula finishing as a bud or bellshape, with the wing membrane pilose. Nielseniella spp. have antennal ascoids with only two branches basally directed and an asymmetric aedeagus.
Previous to this work, only 4 or 5 species had been considered to belong to Feuerborniella : F. oscura (Tonnoir) from the Paleactic region, F. plaumanni (Duckhouse) and F. spathipenis (Duckhouse) both from Brazil, F. malayensis (Satchell) of Malaya ( Vaillant 1974), and apparently another species known only from a single larva from United States ( Vaillant 1983).
The male of Feuerborniella veracruzana sp. nov. is very similar to F. plaumanni , sharing the pattern of head setae patches, the length of the cercopods, general shape of aedeagus tapered to a narrow rounded tip, divergent parameres beneath aedeagus connected to a broad plate above the aedeagus, and the shape of tergite 10. Nevertheless, F. plaumanni can be separated because it has the antennal flagellomeres 11–13 partially fused to each other, the aedeagal apodeme comparatively longer and thinner, the aedeagus much longer than the lateral shafts, and the gonostylus with long apical setae in the male. The female of this species is unknown ( Duckhouse 1968).
Feuerborniella veracruzana sp. nov., can be separated from the rest of the species in the genus: F. oscura has the cercopod as long as epandrium, the apical projection of tergite 10 is short and blunt, the aedeagus is not tapered and the lateral shafts are not curved ectally at the apex in the male, and there is an absence of two strong setae on the genital plate in the female; F. spathipenis has two small projections at the vertex, the aedeagus is spoonlike, and the gonocoxite is nearly as long as the gonostylus in the male.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |