
Phreatobius sanguijuela, Fernandez, Luis, Saucedo, Lisandro J., Carvajal-Vallejos, Fernando M. & Schaefer, Scott A., 2007
|
publication ID |
https://doi.org/ 10.5281/zenodo.179279 |
|
publication LSID |
lsid:zoobank.org:pub:1B412322-3313-4190-A893-8595D6CECE31 |
|
DOI |
https://doi.org/10.5281/zenodo.5679608 |
|
persistent identifier |
https://treatment.plazi.org/id/03B0B034-7745-FF83-FF54-FCD3410AFC46 |
|
treatment provided by |
Plazi |
|
scientific name |
Phreatobius sanguijuela |
| status |
sp. nov. |
Phreatobius sanguijuela View in CoL new species
( Figs. 2–3 View FIGURE 2 View FIGURE 3 )
Holotype. UMSS 1785, 42.1 mm SL, Bolivia, Departamento Santa Cruz, Río Iténez basin, Comunidad de Porvenir, artificial well, 6 m depth near Río Paraguá, 172 m elevation, 13º 59' 14.44'' S, 61º 32' 27.05'' W, collected by L. Saucedo, 1 June 2006. Paratypes. MNKP 5373, 2 specimens: 27.8–32.3 mm SL, same data as holotype.
Diagnosis. Assigned to Phreatobius among siluriform genera based on the possession of the following combination of characters: dorsal and ventral procurrent caudal-fin rays extended rostrally, dorsally reaching a vertical position on dorsal-fin base posterior to dorsal-fin insertion, ventrally continuous with anal fin ( Fig. 2 View FIGURE 2 A); caudal fin round; all anal-fin rays unbranched; mouth prognathous, with jaws displaced dorsally on head; adductor mandibulae muscle hypertrophied, covering most of skull and inserting posteriorly onto first neural spine ( Fig. 2 View FIGURE 2 B); first pectoral-fin ray soft, not spinous; bright red in life. Uniquely diagnosed relative to its congener by the absence of eyes (versus eyes present, minute; Fig. 3 View FIGURE 3 ), presence of 25–34 dorsal procurrent caudal fin rays (versus 42–50); 14–16 ventral procurrent caudal fin rays (versus 22–26); 45–46 vertebrae (versus 54–59).
Description. Morphometric data for the holotype and paratypes presented in Table 1 View TABLE 1 . Body round in cross section from head through dorsal-fin origin, gradually more compressed posteriorly, tapering to caudal fin. Dorsal and ventral trunk profiles nearly straight from head to base of caudal fin. Body depth nearly uniform through middle of anal-fin length, narrowing to caudal-fin base. Dorsal head profile markedly wider than trunk, due to hypertrophied jaw adductor musculature ( Fig. 2 View FIGURE 2 B). Integument of head and body thick, underlain with thick layer of round, adipose bodies individually visible under stereomicroscope; myotomes and intermuscular septa not externally visible.
Head slightly depressed, longer than broad in dorsal view. Snout blunt in dorsal and lateral views. Mouth prognathous, lower jaw extending further anteriorly than upper jaw. Cleft of mouth straight. Lower jaw projected dorsally, its ventral surface markedly convex. Upper jaw strongly depressed, its depth one-third or less than that of lower jaw in lateral view. Mouth as wide as head, its lateral margins strongly curved posteriorly; mouth corner at vertical though anterior third of head length. Upper lips narrow, well defined laterally by fold of integument. Lower lip poorly defined, mostly continuous with ventral surface of head. Eyes absent ( Fig. 2 View FIGURE 2 B). Branchiostegal membranes mostly free, narrowly attached to isthmus anteriorly at midline, with wide and almost free branchial openings; posterior origin of membranes located immediately dorsal to origin of pectoral fin.
Maxillary barbel long and thin, reaching to slightly beyond posterior margin of pectoral-fin base. Outer mental barbel longer than maxillary barbel, reaching pectoral-fin base or posterior third of pectoral fin. Inner mental barbel shortest, when extended parallel to body reaching to point slightly posterior to margin of branchial membrane; its origin slightly anterior to that of lateral mental barbel. All barbels with fine round tips.
Anterior naris prolonged as tube of integument directed anterodorsally, about twice as long as wide, narial opening small, round, located close to upper lip. Posterior naris large, anterior margin crescentic and framed by low rim of elevated integument, located close to base of maxillary barbel but not continuous with its margin. Distance between two posterior nares slightly greater than that between anterior nares.
Cephalic sensory canals comprised of three pores dorsally on anterior snout region near nares. Anterior pair (pore I of Reichel 1927: fig. 3, 305) closely spaced relative to one another along midline, positioned mesially to base of anterior nares. Mid-posterior pair (pore II) located posteromesially to posterior nares. Posterior pair (pore III) located posterolaterally to posterior nares and aligned with maxillary barbel base. Two laterosensory pores visible in lateral view. Anterior-most (pore V) located approximately at median lateral line of body, posterior (pore VI) located slightly laterodorsally. One large pore (IV) located posterolaterally close to end of head. Anterior portion of mandibular canal with two pores (VII, VIII) located ventrally close to barbel and two posterior (IX, X) pores located laterally. Pores of epiphyseal branch absent. Infraorbital canal absent. Laterosensory canal system absent along most of body, restricted anteriorly to short tube with two pores.
Pectoral fin small, base short; four rays total, all soft, first ray unbranched; last ray markedly shorter than anterior rays. Pectoral-fin insertion approximately horizontal. Pelvic fin with four* (n=2) or five (n=1) rays, one or two branched, all projecting beyond fin membrane. Pelvic fin reaching or covering anus, not reaching anal fin. Dorsal-fin margin straight and short, with seven rays (n=3), none, one or three* branched, all soft; fin origin closer to tip of snout than to base of caudal fin, positioned slightly anteriorly to vertical through pelvic fin origin, its posterior insertion approximately at middle of SL. First dorsal-fin pterygiophore inserted posterior to neural spine of vertebra 13. Anal fin straight in profile and long, with 19 (n=2) and 20* (n=1) unbranched rays. Anal-fin origin located slightly posterior to anus and vertical through dorsal fin origin. First anal-fin pterygiophore inserted on centrum 18. Anal-fin continuous posteriorly with ventral procurrent caudal fin rays, fins clearly demarkated by slight indentation along membranous fin margin. Caudal fin margin nearly round, continuous dorsally and ventrally with procurrent rays. Principal caudal-fin rays eight branched plus two unbranched* (n=2) or nine branched plus 2 unbranched (n=1), all soft. Dorsal procurrent caudal-fin rays 25, 31 and 34*. Ventral procurrent caudal fin rays 14, 15, 16*; almost vertically aligned, rays becoming gradually longer posteriorly. Caudal peduncle markedly expanded in area of procurrent rays, like a single extended fin. Vertebrae 45 (n=1) and 46* (n=2). Pleural ribs one or three*.
Pigmentation in alcohol: All specimens nearly whitish, without trace of red coloration characteristic of living specimens. Holotype and largest paratype with pigment concentration along midline of body, forming thin midlateral line ( Fig. 2 View FIGURE 2 A). Tubelike extension of anterior naris white. All barbels white. Pectoral, pelvic, dorsal, anal, and caudal fins, as well as region corresponding to dorsal and ventral procurrent caudal fin rays, lack dark pigment. Coloration in life: Body bright red, slightly translucent, not whitish as in preserved specimens. Outline of vertebral column visible along post-abdominal region. Gut contents evident as dark shadows along ventral translucent region of abdomen.
Etymology. The specific name sanguijuela is the local name for these catfishes in this area of Bolivia, treated here as a noun in apposition to the generic name.
Distribution. Phreatobius sanguijuela is known only from artificial wells of 6 m depth at Comunidad de Porvenir, near Río Paraguá ( Fig. 1 View FIGURE 1 ). The Río Paraguá forms part of the headwaters of the Río Itenéz in eastern Santa Cruz Department, northeastern Bolivia.
Habitat. The artificial wells where P. sanguijuela were collected are separated from the Río Paraguá main channel by approximately 2 km. The wells range in depth from 6 to 12 m with a diameter of approximately one meter, with surface openings covered and sidewalls unfinished. The bottom consists of unconsolidated sand and reddish lime. The water is transparent and water depth is about 1 m. The specimens were captured using a bucket during the dry season (June) and they become much less frequent during the rainy season. Water is used locally for human consumption and the catfish are captured incidentally and killed, often mistaken for parasitic leeches (“sanguijuelas”) due to their bright, blood-red coloration.
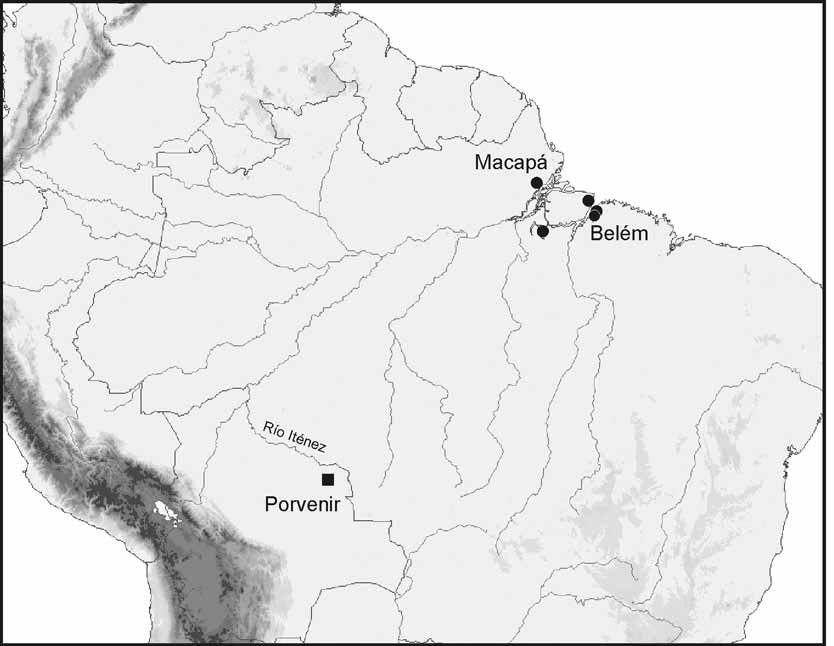

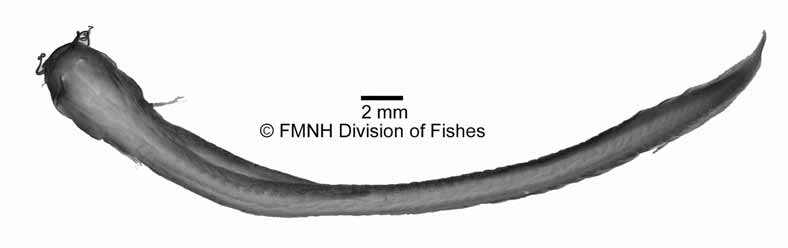
TABLE 1. Morphometric data for holotype and two paratypes of Phreatobius sanguijuela, new species. Standard, total, and head lengths are expressed in millimeters; data 1 – 5 are proportions of standard length; data 6 – 8 are proportions of head length.
| Holotype | Paratype | Paratype | |
|---|---|---|---|
| Standard length (mm) | 42.1 | 27.8 | 32.3 |
| Percentages of SL | |||
| Total length | 1.15 | 1.13 | 1.14 |
| Body depth | 0.10 | 0.09 | 0.09 |
| Caudal length | 0.30 | 0.26 | 0.27 |
| Predorsal length | 0.45 | 0.45 | 0.45 |
| Preanal length | 0.50 | 0.50 | 0.52 |
| Prepelvic length | 0.41 | 0.41 | 0.41 |
| Dorsal-fin base length | 0.10 | 0.09 | 0.09 |
| Head length | 0.17 | 0.16 | 0.16 |
| Head length (mm) | 7.3 | 4.4 | 5.2 |
| Percentages of HL | |||
| Head width | 0.71 | 0.67 | 0.79 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |