
Salina wolcotti Folsom, 1927
|
publication ID |
https://doi.org/10.5281/zenodo.193142 |
|
DOI |
https://doi.org/10.5281/zenodo.5679205 |
|
persistent identifier |
https://treatment.plazi.org/id/03B13638-FF97-FFD7-1DBF-41B626F8F8D9 |
|
treatment provided by |
Plazi |
|
scientific name |
Salina wolcotti Folsom, 1927 |
| status |
|
Salina wolcotti Folsom, 1927: 11 –12, Figs. 61–67 ( Puerto Rico). Wolcott 1933: 241 –242, biology ( Puerto Rico). Essig 1942: 82, mention. Fennah 1947: 64, Fig. 49B, association with corn ( Virgin Islands). Wolcott, 1948: 33, biology ( Puerto Rico). Paclt 1956: 125, mention. Wray 1959: 68, new records (Florida, USA & Costa Rica). Guagliumi 1962: 450 –452, on sugarcane ( Venezuela). Martin & Gregory 1962: 297, biology ( Puerto Rico). Metcalfe 1964: 24, association with sugarcane ( Jamaica). Mari Mutt 1978: 5, repository of syntypes. Massoud & Thibaud, 1980: 604, mention. Mari Mutt 1982: 32, catalog of Collembola of Puerto Rico. Mari Mutt 1987b: 406, Fig. 57, taxonomic status. Escher & Lounibos, 1993: 483, association with Pistia stratiotes (Florida) View in CoL . Carithers 1997: 148, as prey of hymenopteran Microstigmus comes View in CoL ( Costa Rica). Heckman 2000: 287, Fig. 1447, key to Salina species. Palacios- Vargas 2000: 5, catalog ( México). Palacios-Vargas 2003: 221, 223, mention ( México).
Material Examined: Puerto Rico. Río Piedras; 23 February 1920; G. N. Wolcott, coll.; 1 slide-mounted syntype. Bayamón; 5 May (year not included); G. N. Wolcott, coll.; 1 slide-mounted syntype. Toro Negro Commonwealth Forest, N18° 10.197, W66° 29.536; beating grasses and Impatiens along road (PR–143) and along El Bolo trail, near forest administration building; 29 July 1999; 9 individuals on slides
Description: The condition of the slide-mounted syntype from Bayamón is such that none of the diagnostic characters are visible. The syntype from Río Piedras clearly shows 2 macrosetae on Ant. 1, 5 setae along the cephalic groove and 3, 4, 2, 2 macrosetae on Th. 2-Abd. 2. Only one prelabral seta is visible. Other potentially diagnostic characters included in Table 1 View TABLE 1 (e.g., posterior setae on Abd. 4 and posterior setae on collophore) cannot be discerned on the cleared specimen. Therefore, the description that follows is based on the specimen collected in Toro Negro.
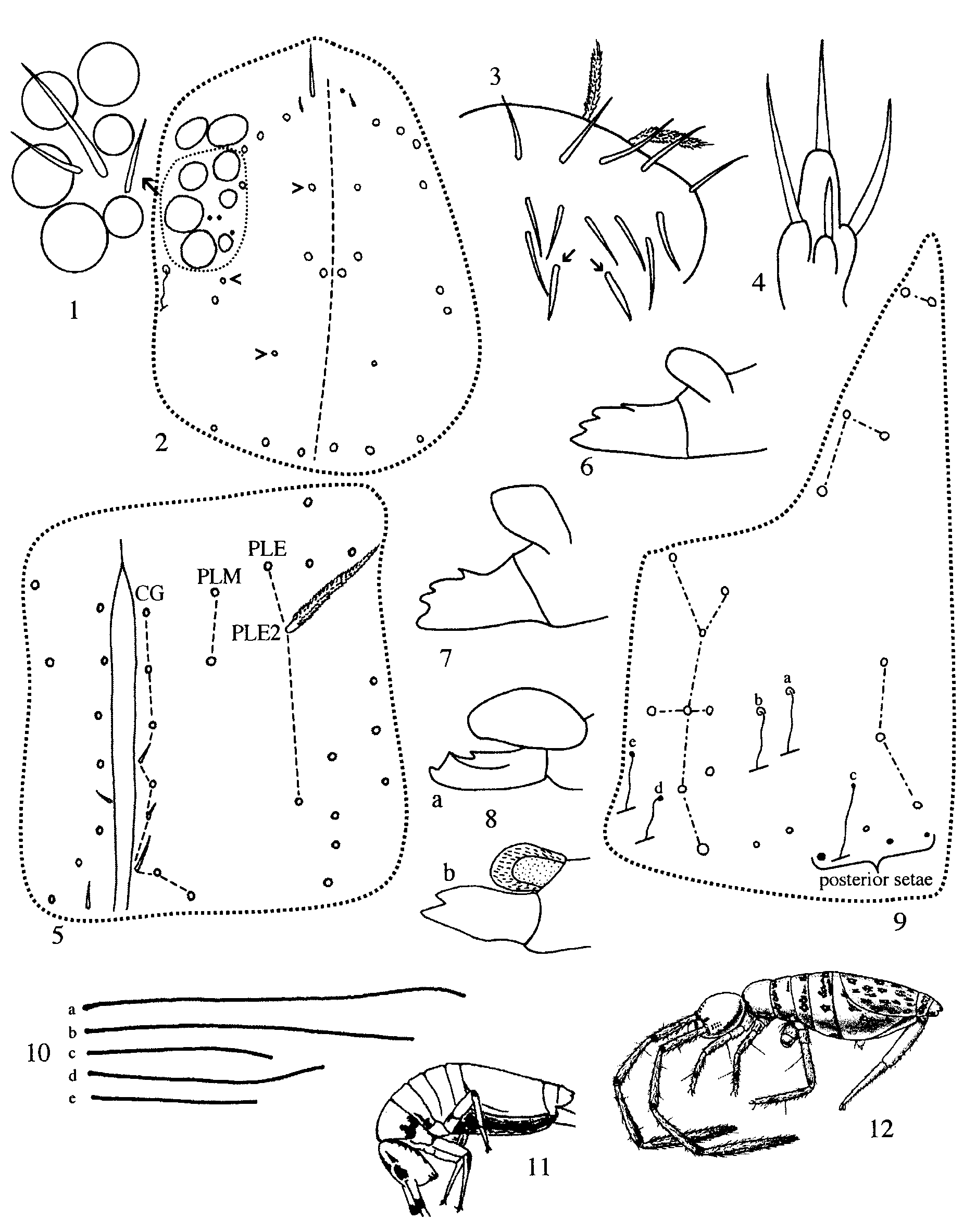
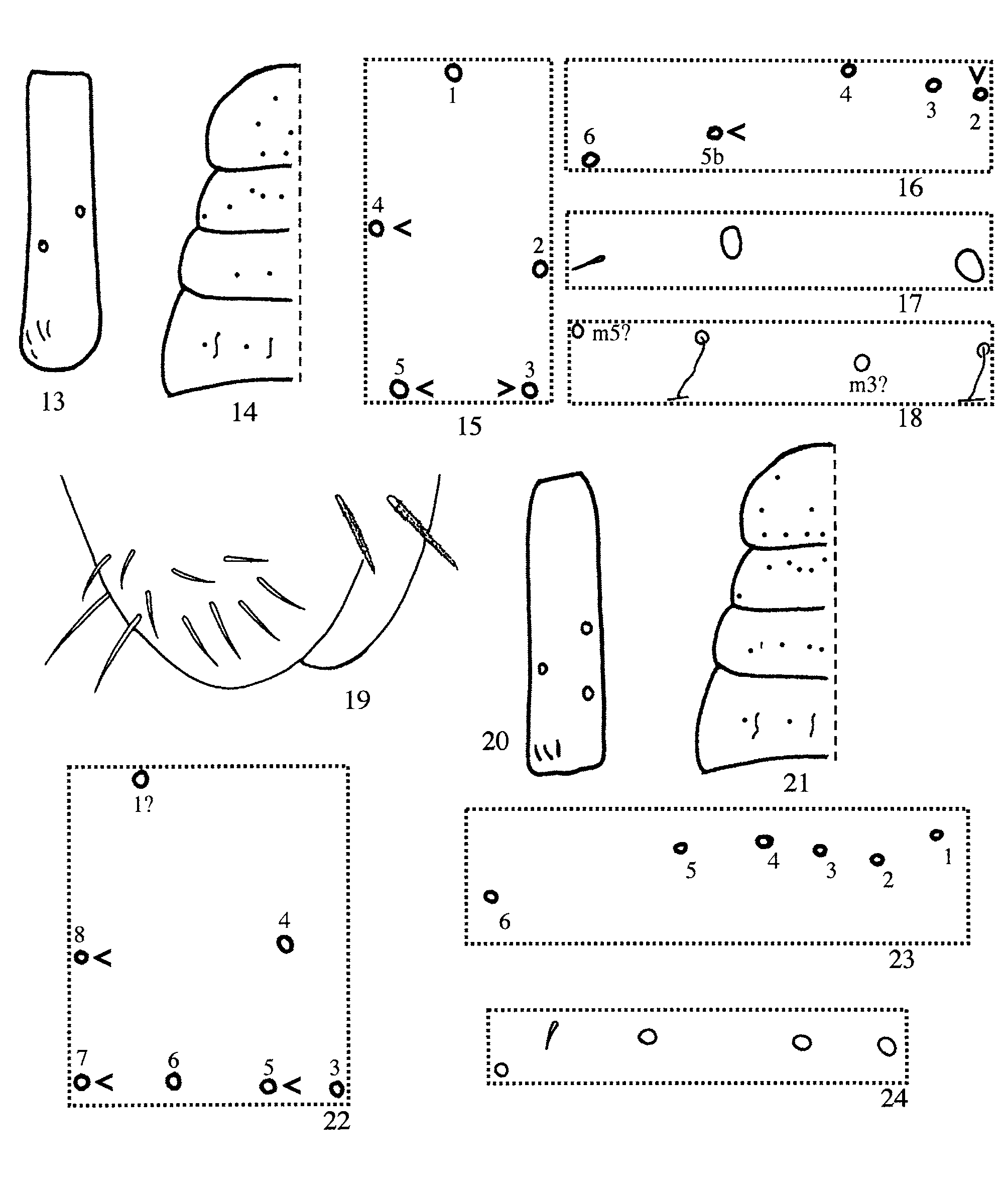
Length to 1.4 mm (smallest individual= 1.1 mm; average=1.2 mm; n=8). Original colour pattern described by Folsom (1927) and shown in Fig. 12 View FIGURES 1 – 12. 1 – 4 seen only in four poorly preserved syntypes (including the slidemounted syntype from Bayamón listed above). Freshly collected specimens background colour yellowish green, which fades to white in individuals preserved in alcohol for a long time (> ten years). Most individuals from Toro Negro and the syntype from Río Piedras without dorsal blue markings, except for a blue dot between antennae; some specimens with blue pigment distributed as described for populations of S. tristani from Puerto Rico (cf. Mari Mutt 1987b, Fig. 53) and S. ventricolor from Cuba ( Fig. 11 View FIGURES 1 – 12. 1 – 4 ), forming a narrow band along the anterior and lateral margins of Th. 2 which extends to margins of Th. 3 or Abd. 1; blue pigment forming discreet dots distributed on postero-ventral field of head and on sternal plate of abdomen that accommodates folded furcula. Antennae amber, tip of each segment with dark blue ring; legs amber, lighter than antennae. Collophore and furcula white. Ant. 1 with 2 dorsal macrosetae ( Fig. 13 View FIGURES 13 – 24 ). Prelabral setae 2. Postlabial column CG with 5 (3–6) ciliate acuminate setae, most individuals (6 of 9) with asymmetric number of setae. Tergal chaetotaxy of Th. 2–Abd. 2 as 3(3–5), 4(2–5), 2(2–3), 2 ( Figs. 14–18 View FIGURES 13 – 24 ): Th. 2 setae numbered 3–5 in Figure 15 View FIGURES 13 – 24 sometimes reduced or absent, seta 3 most often present; Th. 3 setae 2 and 5b absent in 2 and 1 individuals, respectively; Abd. 4 with 3 posterior setae ( Fig. 9 View FIGURES 1 – 12. 1 – 4 ). Trochanteral organ with 14 (11–18) setae. Unguis with 2 distal unpaired teeth. Collophore anterior face with 3 microsetae and 3 distal macrosetae; lateral setae ( Fig. 19 View FIGURES 13 – 24 ) 10–11; posterior face with 1+1 microsetae. Proportions of dens/manubrium≈ 1, proportion of dental vesicle/mucro≈ 1. Mucro with dorso-basal denticle ( Fig. 8 View FIGURES 1 – 12. 1 – 4 ).
Remarks: Salina wolcotti differs from S. bidentata in the number of dorsal macrosetae on Ant. 1, Th. 2– Abd.1 (2,3,4,2, respectively in wolcotti ; 3,6, 6,4 in bidentata— see Table 1 View TABLE 1 for variation range) and number of posterior setae on collophore (1+ 1 in wolcotti ; 2+ 2 in bidentata ). Salina thibaudi n. sp. has identical amoeboid colour pattern as illustrated for S. wolcotti by Folsom (1927) but it differs in practically all important chaetotaxic characters, including the number of prelabral setae ( 2 in S. wolcotti , 4 in S. thibaudi ), the number of macrosetae on Ant. 1, Th. 2–Abd. 1, number of posterior setae on Abd. 4, and posterior setae of collophore as shown in Table 1 View TABLE 1 . Salina wolcotti differs from S. ventricolor in the number of macrosetae on Th. 2 ( 3 in wolcotti , 4 in ventricolor ), pattern of macrosetae on Th. 3 (seta 5b present and 5 absent in wolcotti whereas 5b absent and 5 present in ventricolor ), number of anterior setae on collophore ( 6 in S. wolcotti ; 7 in ventricolor ) and possibly in head chaetotaxy. However, these differences may be illusory since no variation in number of setae was reported for S. ventricolor . With the exception of chaetotaxy on Th. 3 and anterior face of collophore, all characters described for S. ventricolor fall within the range of variation observed in the nine individuals of S. wolcotti studied from Puerto Rico. Closer examination of S. ventricolor populations may show this form to be a junior synonym of S. wolcotti . Salina wolcotti differs from S. beta in colour pattern, number of macrosetae on Th. 3–Abd. 1, and number of posterior setae on Abd. 4.
The syntypic series of S. wolcotti comprises specimens collected at unspecified localities in Punta Cangrejos (Carolina), Río Piedras, Bayamón, Isabela and Guánica. The syntypes deposited at the Illinois Natural History Survey were collected at Río Piedras and Bayamón, but the accompanying labels do not provide additional information about the localities. All the general localities mentioned above, except Carolina, were sampled in the late 1970 or early 1980 by Mari Mutt (1987b) or the author, but the only species collected at these localities was S. tristani . Based on the redescription and circumscription presented above, S. wolcotti appears to be endemic to Puerto Rico, with a single population remnant in the central mountain region of the island. Records of S. wolcotti from Florida, USA and Costa Rica ( Wray 1959) are probably referable to S. bidentata or S. thibaudi , n. sp. (see below), but I have not studied that material. Reports of S. wolcotti from other localities around the Caribbean basin need to be confirmed using the characters described above.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Salina wolcotti Folsom, 1927
| Soto-Adames, Felipe N. 2010 |
Salina wolcotti
| Palacios-Vargas 2003: 221 |
| Heckman 2000: 287 |
| Vargas 2000: 5 |
| Carithers 1997: 148 |
| Escher 1993: 483 |
| Mari 1987: 406 |
| Mari 1982: 32 |
| Massoud 1980: 604 |
| Mari 1978: 5 |
| Metcalfe 1964: 24 |
| Guagliumi 1962: 450 |
| Martin 1962: 297 |
| Wray 1959: 68 |
| Paclt 1956: 125 |
| Wolcott 1948: 33 |
| Essig 1942: 82 |
| Wolcott 1933: 241 |
| Folsom 1927: 11 |