
Dasypoda michezi Radchenko, 2017
|
publication ID |
https://doi.org/10.11646/zootaxa.4350.1.10 |
|
publication LSID |
lsid:zoobank.org:pub:811A55CA-CC41-4C11-81FC-BB7DD0E47CC9 |
|
DOI |
https://doi.org/10.5281/zenodo.6035210 |
|
persistent identifier |
https://treatment.plazi.org/id/03B1F643-FFE9-E77B-FF6C-21AA68487729 |
|
treatment provided by |
Plazi |
|
scientific name |
Dasypoda michezi Radchenko |
| status |
sp. nov. |
Dasypoda michezi Radchenko View in CoL , sp. n.
Type material: Holotype ♂ with the printed label: “ Dasypoda ♂, Almograve (Port[ugal].) [37.6524, -8.7960], 21- V-2012, A.Livory & R.Coulomb leg.”.
Paratype: 1 ♂ with the handwritten label: “ Dasypoda ♂, Vila Nova de Milfontes (P[ortugal]) [37.723, -8.788], 20-V-2012, A.Livory u R.Coulomb leg.”. Both specimens are stored in the collection of Naturalis Biodiversity Center, Leiden, the Netherlands ( RMNH).
Etymology. Named after Denis Michez, an authority on Dasypoda and other bees.
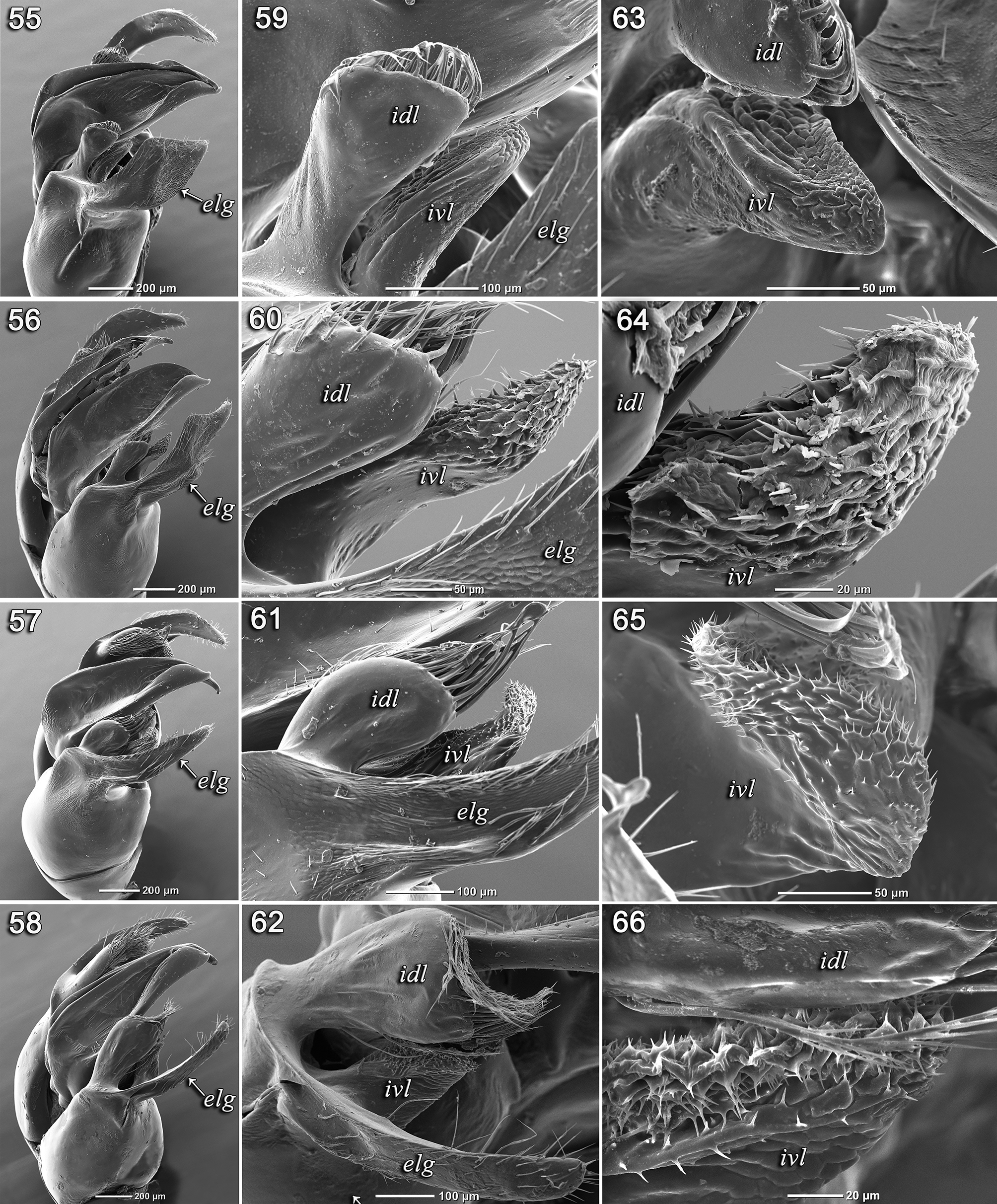
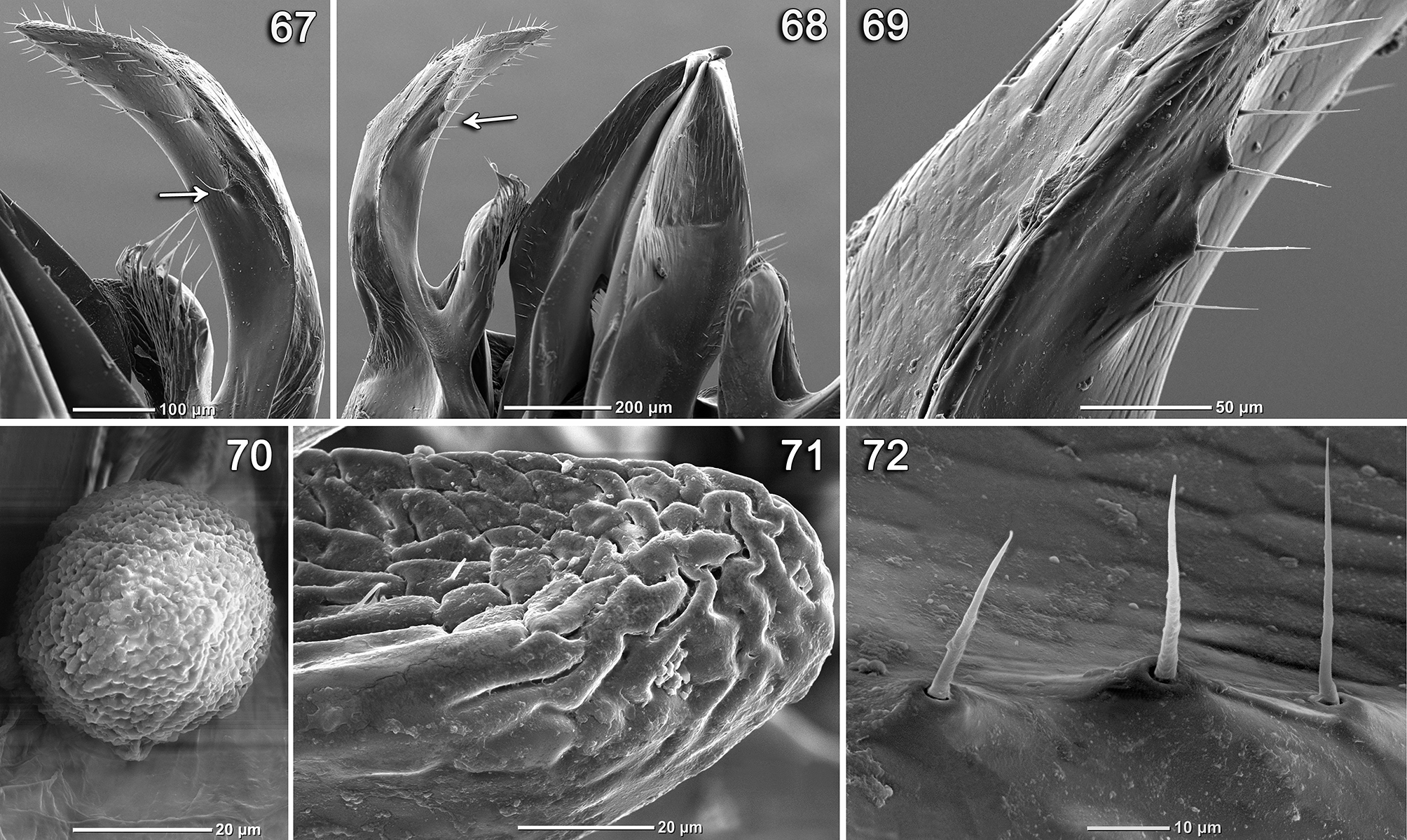
Diagnosis. Dasypoda michezi sp. nov. may be regarded a representative of the subgenus Heterodasypoda due to the possession of all its diagnostic features, namely the shape and structure of maxillary palpi, galea ( Fig. 7 View FIGURE 7 – 14 ), malar space, nervulus ( Fig. 1 View FIGURE 1 – 6 ), S6–8 ( Figs 23, 27, 31 View FIGURE 23 – 34 , 35, 39 View FIGURE 35 – 42 ), genitalia ( Figs 43, 47, 51 View FIGURE 43 – 54 , 55 View FIGURE 55 – 66 ), and gonostylus ( Figs 59, 63 View FIGURE 55 – 66 , 67, 68 View FIGURE 67 – 72 ). The new species differs from other species of this subgenus by having: apical projection of S6 apically deeply emarginated in the form of wide triangle ( Fig. 23 View FIGURE 23 – 34 ), apicolateral processes of S7 with triangle shaped distal expansion ( Fig. 28 View FIGURE 23 – 34 ), inner ventral lobe of gonostylus with only few setae ( Figs 63 View FIGURE 55 – 66 , 71 View FIGURE 67 – 72 ), distal half of external lobe of gonostylus significantly widened and convex in middle of outer surface, diamond-shaped ( Figs 43, 47, 51 View FIGURE 43 – 54 , 55 View FIGURE 55 – 66 ) and its inner surface with several hairs, each of which inserted into crater-like raised alveoli ( Figs 67–69, 72 View FIGURE 67 – 72 ).
Description. Male: Holotype. Body length (vertex to T7): 11.8 mm (paratype: 12.0 mm). Mesosoma: W (between tegulae) = 3.0 mm (same in paratype). Head: L = 2.9 mm (paratype: 3.1 mm), W = 3.3 mm (paratype: 3.5 mm);
Body black, head slightly wider than long ( Figs 3, 4 View FIGURE 1 – 6 ), clypeus, densely punctured by oblique, downward punctures. Lateral edges of clypeus with impunctate bands on sides and apex of clypeus with narrow depressed band ( Fig. 4 View FIGURE 1 – 6 ). Face below antennal toruli with relatively wide (equal to the width of scapus), smooth, almost impunctate band reaching base of clypeus. Malar space narrow, 0.15× as long as broad, in 2× shorter than pedicel. Body mostly covered with light yellow-grey hairs ( Figs 1–6 View FIGURE 1 – 6 ); head also with such hairs except upper part of face between compound eyes and vertex covered with sparse brown hairs ( Figs 3, 4 View FIGURE 1 – 6 ). Back of head and thorax with long, semi-erect hairs. Wings slightly infuscate, notably in distal half. Legs with long greyish-yellow pubescence, apart from dense short dark-yellow pubescence of inner side of metabasitarsus, and very short grey hairs of 2–5th tarsomeres with a sparse sheaf of long hairs on apices of 2–4th tarsi. T1–2 covered by light, long erect hairs, T3–5 with semi-erect hairs and T6–7 with appressed hairs (T5–6 with pale pubescence only on the margin and sides), each hair rising from an oblique backward-directed puncture. Distances between puncture about 2–4× as long as diameter of puncture. Pubescence sparse and revealing surface of tergum. T2–6 with very short, thin branched dark-brown hairs situated in areas between long hairs (length of lateral branches equal to 1/2–2/3 of total hair length). T5–6 with basal parts bearing only dark-brown hairs with small admixture of light hairs on sides. Basal parts of sterna densely punctate, mainly bearing dark brown hairs. Central part of S5–6 with larger sparse punctures. S3–5 with apical bands of appressed dark-brown hair, with longer light yellow-grey hairs on sides of sterna ( Fig. 6 View FIGURE 1 – 6 ). Apical part of S6 with wide and deeply emarginated outgrowth ( Fig. 23 View FIGURE 23 – 34 ); distal part of latero-apical processes of S7 triangularly expanded ( Fig. 28 View FIGURE 23 – 34 ); apex of S8 posterodorsal part with transverse carina slightly notched in center ( Fig. 35 View FIGURE 35 – 42 ; carina shown by white arrow). Inner ventral lobe of gonostylus almost bare on apex ( Figs 63 View FIGURE 55 – 66 , 71 View FIGURE 67 – 72 ); distal half of external lobe of gonostylus significantly widened and convex on its outer surface, diamond-shaped ( Figs 43, 47, 51 View FIGURE 43 – 54 , 55 View FIGURE 55 – 66 ), its inner surface with several hairs, each inserted into crater-like raised alveoli ( Figs 67–69, 72 View FIGURE 67 – 72 ).
Female: unknown.
Distribution. South-western Portugal ( Fig. 73).
Floral visitation. Pollen grains of a Cistus plant ( Cistaceae ) were found on the body of the paratype ( Fig. 70 View FIGURE 67 – 72 ). Other representatives of this subgenus, in particular D. morotei and D. pyrotrichia , are also associated with Cistaceae ( Michez et al. 2004b, 2008; Ruiz 2013; Özbek 2014), whereas D. albimana is mostly found on Asteraceae , but also recorded on Cistaceae , Lamiaceae and Rosaceae ( Michez et al. 2003, 2004b; Ruiz 2013).
Flight period. May. All other Heterodasypoda species also fly in late spring to summer: D. albimana —from May to July ( Michez et al. 2003); D. morotei —from May to August, with some sporadic records in April and September and one—later in November (Ornosa & Ortiz-Sánchez 1998, 2004); D. pyrotrichia —from mid May to late August ( Grace 2010).
Comparative remarks. Dasypoda michezi sp. nov. is most likely to be confused with D. albimana . Both species have similar body length and size of head, approximately same surface punctation of different body parts and colour of pubescence. However, D. michezi sp. nov. is characterized by the shorter first flagellar segment ( Figs 15, 19 View FIGURE 15 – 22 ) and differs from all other species of the subgenus in the ratio of different flagellar segments ( Figs 17–18, 21–22 View FIGURE 15 – 22 ; Tabl. 1).
Flagellomeres 3–8 of D. michezi sp. nov. are cylindrical ( Fig. 15 View FIGURE 15 – 22 ), the same as in D. albimana ( Fig. 16 View FIGURE 15 – 22 ) and D. pyrotrichia ( Fig. 18 View FIGURE 15 – 22 ), in contrast to D. morotei , in which they are slightly curved ( Fig. 17 View FIGURE 15 – 22 ). The clypeus of D. michezi sp. nov. ( Figs 3, 4 View FIGURE 1 – 6 ) lacks an impunctate median band (this feature is almost invisible when pubescence is undamaged), likewise in D. albimana . Also, the apex of the clypeus is straight in D. michezi sp. nov. (the clypeus of D. morotei and D. pyrotrichia has a more or less distinct, impunctate median band; also it the apex is emarginated in D. morotei ). The glossa of D. michezi sp. nov. ( Fig. 11 View FIGURE 7 – 14 ) is very narrow and long: 1.5× longer than in D. albimana ( Fig. 12 View FIGURE 7 – 14 ) and D. morotei ( Fig. 13 View FIGURE 7 – 14 ), but has same length as in D. pyrotrichia ( Fig. 14 View FIGURE 7 – 14 ). The most noticeable differences concerns the ratio of the glossa width at base to its length: this proportion is about 1: 6.7 in D. michezi sp. nov., 1: 4.6 in D. pyrotrichia , 1: 3 in D. albimana and 1: 2.3 in D. morotei ( Figs 11–14 View FIGURE 7 – 14 ). The surface of the galea of D. michezi sp. nov. ( Fig. 7 View FIGURE 7 – 14 ) is clearly structured with many small tubercles merged into sinuous lines in many places, and is less shiny than in D. albimana and D. pyrotrichia ( Figs 8, 10 View FIGURE 7 – 14 ). This surface is completely covered with separate small tubercles in D. morotei ( Fig. 9 View FIGURE 7 – 14 ).
Species Flagomere L: W (µm) Flagomere L: W ratio Flagomeres length ratio
1st* 2nd 1st* 2nd 3rd ... 9th 1st: 2nd
Dasypoda michezi sp. nov. (n = 2) 412/290 320/295 1.41 1.08 1.1÷1.2 1.28
D. albimana (n = 4) 475/257 268/263 1.85 1.02 1.1÷1.2 1.77
D. morotei (n = 2) 358/293 398/284 1.22 1.40 1.4÷1.5 0.90
D. pyrotrichia (n = 3) 450/290 370/296 1.55 1.25 1.4÷1.6 1.22
*—width on apex.
The main significant differences between D. michezi sp. nov. and the other three species of Heterodasypoda concern the structure of S6–8 of males. The outgrowth on apical part of S6 is significantly expanded at apex (ratio between width of outgrowth at base to its width at apex is 1: 1.25), apically deeply emarginated in the form of wide triangle ( Fig. 23 View FIGURE 23 – 34 ) in D. michezi sp. nov., unlike other species in which such outgrowth on apex is not expanded (in D. albimana and D. morotei ; Figs 24, 25 View FIGURE 23 – 34 ) or almost undeveloped (in D. pyrotrichia ; Fig. 26 View FIGURE 23 – 34 ). The new species also differs in the structure of latero-apical process at S7 ( Figs 27–30 View FIGURE 23 – 34 ). S8 of D. michezi sp. nov. is chunkier ( Figs 31 View FIGURE 23 – 34 , 39 View FIGURE 35 – 42 ), and superficially appears similar to D. albimana ( Figs 32 View FIGURE 23 – 34 , 40 View FIGURE 35 – 42 ), but significantly differs from that of D. morotei , which has undulating depressions on the sides of basal part of this segment ( Figs 33 View FIGURE 23 – 34 , 41 View FIGURE 35 – 42 ; shown by black arrows), and D. pyrotrichia is possessing more extended sides on the basal part of this segment ( Figs 34 View FIGURE 23 – 34 , 42 View FIGURE 35 – 42 ). The transverse carina situated anteriorly to the apex of the inner part of S8 has a small notch in the center in D. michezi sp. nov. ( Fig. 35 View FIGURE 35 – 42 ), (cf. D. albimana and D. morotei ( Figs 36, 37 View FIGURE 35 – 42 ) in which it is more deeply cut in or even completely bilobed in D. pyrotrichia ( Fig. 38 View FIGURE 35 – 42 )).
The inner ventral lobe of the gonostylus is almost bare in D. michezi sp. nov., with a few thin and short setae ( Figs 63 View FIGURE 55 – 66 , 71 View FIGURE 67 – 72 ), unlike other species, in which this lobe is covered with numerous, more or less long and pointed setae ( Figs 64–66 View FIGURE 55 – 66 ). The distal half of external lobes of the gonostylus of D. michezi sp. nov. is considerably widened and convex in the middle of the outer surface, diamond-shaped ( Figs 43, 47, 51 View FIGURE 43 – 54 , 55 View FIGURE 55 – 66 ). Unlike the new species, these lobes are narrow or more strongly elongated toward apex and widely concave in the middle of outer surface in other discussed species ( Figs 44–46, 48–50, 52–54 View FIGURE 43 – 54 , 56–58 View FIGURE 55 – 66 ). The inner medial surface of the outer lateral lobe of gonostylus is bearing several hairs, each of which is inserted into the crater-like raised alveoli ( Figs 67–69, 72 View FIGURE 67 – 72 ) in D. michezi sp. nov.; no such hairs are found in other species of Heterodasypoda .






| RMNH |
National Museum of Natural History, Naturalis |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Apoidea |
|
Family |
|
|
Genus |
|
|
SubGenus |
Heterodasypoda |