
Microtus agrestis ( Linnaeus, 1761 )
|
publication ID |
https://doi.org/10.1093/mspecies/sex003 |
|
publication LSID |
lsid:zoobank.org:pub:71F30AF5-D653-4941-8E6B-C30B571EBFCA |
|
DOI |
https://doi.org/10.5281/zenodo.4593825 |
|
persistent identifier |
https://treatment.plazi.org/id/03B3878F-B659-FFB7-76B8-FB46FAC23A1E |
|
treatment provided by |
Felipe |
|
scientific name |
Microtus agrestis ( Linnaeus, 1761 ) |
| status |
|
Microtus agrestis ( Linnaeus, 1761)
Field Vole
Mus agrestis Linnaeus, 1761:11 . Type locality “Svecia [= Sweden],” Uppsala.
Mus gregarius Linnaeus, 1766:84 . Type locality “ Germania, Svecia [= Germany and Sweden],” East Prussia.
Mus arvalis nigricans Kerr, 1792:239 . Replacement name for Mus agrestis Linnaeus, 1761 .
Arvicola hirta Bellamy, 1839:373 . Type locality “Yealmpton,” Devonshire, England.
Arv. [ icola] Bailloni de Sélys Longchamps, 1841:225. Type locality “Abita il Nord della Francia e della Svizzera [=Inhabits the North of France and Switzerland.”
Arvicola neglecta Jenyns, 1841:270 . Type locality “Megarnie Castle in Perthshire,” Scotland.
Arvicola levernedii Crespon, 1844:73 . Type locality “entre St-Gilles et Aiguesmortes [= between St. Gilles and Aiguesmortes],” Gard, France.
Lemmus insularis Nilsson, 1844:34 . Type locality “Ostgötha, skärgård,” Sweden.
A. [ rvicola] britannicus de Sélys Longchamps, 1847:307. Type locality “L’Angleterre et l’Écosse [= England and Scotland].”
Arvicola rozianus Bocage, 1865:7 . Type locality “ Coimbra, no sitio da Geria,[= Coimbra, county of Geria],” Portugal.
Arvicola agrestis var. nigra Fatio, 1869:241 . Type locality “près d’Engstlen [= vicinity of Engstlen],” Berne, Switzerland.
[ Arvicola agrestis var.] rufa Fatio, 1900:472. Type locality “dans les Alpes savoisiennes, frontière du Valais, et dans les Alpes vaudoises (aux plans de Frenières),” Geneva, Switzerland.
Arvicola agrestis angustifrons Fatio, 1905:191 . Type locality “près de Zermatt, entre 1620 et 1700 m. s/m., en Valais [= near Zermatt between 1,620 and 1,700 meters in Valais],” Switzerland.
Arvicola agrestis latifrons Fatio, 1905:194 . Type locality “sur quelques points en Suisse, dans les bois de Veyrier, près de Geneve; entre autres et dans les environs de Lucerne [= at some points in Switzerland, in the woods of Veyrier near Geneva; including in and around Lucerne].”
Microtus agrestis exsul Miller, 1908:201 . Type locality “North Uist, Hebrides,” Scotland.
Microtus agrestis insul Lydekker, 1909:74 . Incorrect subsequent spelling of Lemmus insularis Nilsson, 1844 .
Microtus agrestis mongol Thomas, 1911:759 . Type locality “Kemtchik Valley, Tannu-ola Mts., N.W. Mongolia.”
Microtus agrestis arcturus Thomas, 1912:398 . Type locality “Barlik Mts. S., N.W. Dzungaria,” China.
Microtus agrestis mial Barrett-Hamilton and Hinton, 1913a:364 . Type locality “Eigg, Inner Hebrides,” Scotland.
Microtus agrestis luch Barrett-Hamilton and Hinton, 1913a:366 . Type locality “Muck, Inner Hebrides,” Scotland.
Microtus agrestis macgillivrayii Barrett-Hamilton and Hinton, 1913b:831 . Type locality “Islay,” Inner Hebrides, Scotland.
Microtus agrestis fiona Montagu, 1922:940 . Type locality “Gigha,” Inner Hebrides, Scotland.
Microtus agrestis punctus Montagu, 1923:868 . Type locality “ Bled, Slovenia.”
Microtus hirtus orioecus Cabrera, 1924:8 . Type locality “Molins, Montseny, Prov. Gerona,” Spain.
Microtus agrestis pannonicus Ehik, 1924:76 . Type locality “Orman, near Komarviros County Zala, Hungary.”
Microtus agrestis tridentinus Dal Piaz, 1924:10 . Type locality “Brennero, Alto Adige, 1400 m s/m,” Italy.
Microtus agrestis estiae Reinwaldt, 1927:13 . Type locality “Inseln Villsandi und Abruka, [= Villsandi and Abruka Islands],” Estonia.
Microtus agrestis Wettsteini Ehik, 1928:197 . Type locality “Ob. Trixen, Karintia [= Carinthia],” Austria.
Microtus agrestis ognevi Skalon, 1935:11 . Type locality “Церковенское на р. Таз [=Tserkovenskoe on Taz River],” northwestern Siberia.
Microtus agrestis pallida Melander, 1938:74 . Type locality “Смоленская обл, Ельнинский р- н. [= Elninsky department of Smolensk province, near vill. Miloje],” Russia.
Microtus agrestis argyropoli Ognev, 1944:179 . Type locality “South Urals, Valley of Inzer River,” Russia.
Microtus agrestis scaloni Heptner, 1948:710 . Replacement name for Microtus agrestis ognevi Skalon, 1935 .
Microtus agrestis argyropuli Ognev, 1950 : Replacement name for Microtus agrestis argyropoli Ognev, 1944 .
Microtus agrestis carinthiacus Kretzoi, 1958:57 . Replacement name for M. a. wettsteini Ehik, 1928.
Microtus agrestis enez-groezi Heim de Balsac and de Beaufort, 1966:638 . Type locality “Groix (= Morbihan),” France.
Microtus agrestis armoricanus Heim de Balsac and de Beaufort, 1966:638 . Type locality “environs de Quimper (= Finistère),” France.
CONTEXT AND CONTENT. Order Rodentia , suborder Myomorpha , superfamily Muroidea , family Cricetidae , subfamilyArvicolinae, genus Microtus , subgenus Microtus . There remains some uncertainty concerning valid subspecies. Musser and Carleton (2005) list the above 39 named forms of Microtus agrestis without reference to subspecies.
NOMENCLATURAL NOTES. Microtus agrestis and the Nearctic M. pennsylvanicus are similar morphologically ( Klimkiewicz 1970). However, this similarity represents convergence, not a phylogenetic relationship (Krapp and Niethammer 1982; Musser and Carleton 2005). Phylogenetic analyses based both on karyotypes ( Modi 1987) and DNA sequences of the entire cytochrome -b gene (Conroy and Cook 2000; Jaarola et al. 2004) clearly show that M. agrestis and M. pennsylvanicus are separate species. The European populations of M. agrestis are differentiated as 3 evolutionary units that may represent cryptic species ( Kryštufek et al. 2008; Paupério et al. 2012).
The generic name Microtus is a combination of 2 Greek words, micros for small and ous for ear, and the specific epithet agrestis is a Latin word meaning “of the field.” Vernacular names assigned to M. agrestis include common field vole, grass mouse, short-tailed vole, dark vole, blackish meadow mouse, and shorttailed field mouse (Alibhai and Gipps 1991; MacDonald and Barrett 1993; Kryštufek et al. 2008).
DIAGNOSIS
Juvenile Microtus agrestis , the common vole ( M. arvalis ), the root vole ( M. oeconomus ), and the East European vole ( M. levis —formerly M. rossiaermeridonalis ) are virtually indistinguishable by gross morphology ( Nekrutenko et al. 2000). Representational difference analysis was used to isolate species-specific markers—Mag3 for M. agrestis and Moe1 for M. oeconomus ( Nekrutenko et al. 2000) . Using these sequences, species-specific polymerase chain reaction (PCR) primers produced unique amplification products that were able to distinguish between these 2 species.
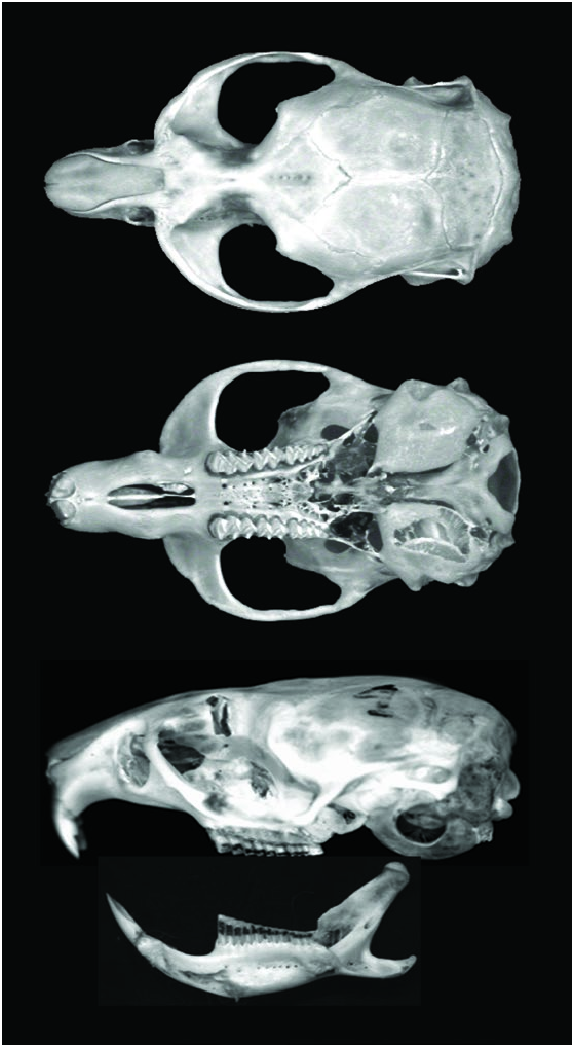
Microtus agrestis ( Fig. 1 View Fig ) can be characterized by its pelage, rather long, shaggy, grayish brown, often tinged with russet and by its bicolored tail with dark dorsal fur. Tail length is about one-third the length of head and body. Only M. duodecimcostatus and M. lusitanicus have shorter tail lengths. These characters of M. agrestis combined with some or all (depending on which comparison is being made) of the following are diagnostic of the species: skull moderately broad, with width> one-half the length; width of interorbital constriction> 3.3 mm; M2 with 3 inner ridges ≤ 5 fields: an anterior loop, 2 outer and 1 inner closed triangles, a character absent in other European vole species, and a 5th field in a postero-internal position; M3 with 5 fields: an anterior loop, 3 closed triangles, and a long terminal loop rarely simplex; m1 with ≥ 7 fields and 4 outer ridges on the outer side; m3 with 3 transverse fields; length of upper molar toothrow never> 7 mm; foramen mandibulare usually lying over protuberance of lower incisor ( Miller 1912; Ellerman 1940; Reichstein 1959; Saint-Girons 1973; Corbet 1978; Krapp and Niethammer 1982).
Microtus agrestis is very similar to M. arvalis , but the latter can be distinguished by its pelage that is more yellowishbrown and its slightly smaller size (body length of M. arvalis 90–120 mm, body weight 14–40 g—Krapp and Niethammer 1982; MacDonald and Barrett 1993). Hairs inserted near the base of the ear are shorter in M. arvalis than M. agrestis ( Dienske 1969) . The asynaptic X and Y chromosomes of M. agrestis contain additional, nonobligatory heterochromatic blocks not found in another species of asynaptic vole, the Mediterranean pine vole ( M. duodecimcostatus ) as addressed by Borodin et al. (1995).
GENERAL CHARACTERS
General form of Microtus agrestis is robust with a large head (broad posteriorly), short and rounded ears, small eyes with diameter of 3.1–3.8 mm (Krapp and Niethammer 1982), and blunt snout. Mouth is small with upper incisors projecting very slightly. Tail is scantily covered with hair and has as many as 20 visible rings per cm near middle ( Miller 1912; Krapp and Niethammer 1982). Ears are mostly covered by fur; meatal lobe (length) is frequently> l mm though < 2 mm ( Dienske 1969). General color of dorsum varies from dark gray-brown to chestnut-brown, with underparts pure gray or pale brown (Corbet and Harris 1991; MacDonald and Barrett 1993; Mathias 1999). Juveniles are darker than adults. M. agrestis has 2 pairs of pectoral and 2 pairs of inguinal mammae. Hind feet have 6 plantar pads, whereas forefeet have 5 palmar pads and inner digit is reduced to a minute tubercle ( Miller 1912).
Dorsal profile of skull is moderately convex. Condyles are visible in dorsal view due to forward projection of occiput ( Fig. 2 View Fig ). In older animals, outer edges of parietals have welldeveloped longitudinal ridges that are continuous posteriorly with lambdoidal crest and which come together abruptly anteriorly to join interorbital ridge ( Miller 1912). Auditory bullae are large and evenly inflated. Interorbital region is narrow, subcylindrical, and has distinct temporal ridges, which can unite to form a definite median crest extending well forward to nasals (Magalhães and Madureira 1980). Nasals are broad anteriorly and abruptly narrow at middle; posterior end is pointed, blunt, or angular. Incisive foramen is long and narrow, extending approximately to molar alveoli. Well-developed postorbital processes are present (Magalhães and Madureira 1980). Posterior edge of foramen mandibulare is about 0.5 mm from posterior edge of mandible ( Dienske 1969; Fedyk and Ruprecht 1971). Mandible is robust and masseteric ridge is well developed.
Lower incisors are much less strongly curved than upper incisors. Molar teeth are open-rooted throughout life, decreasing in size posteriorly. A postero-internal loop is present in M1, similar to that in M2; this is a frequent character in some populations (morphotype exsul — Miller 1912; Ognev 1950; Niethammer 1964; Reichstein and Reise 1965; Reichstein 1966; Meylan 1967; Almaça 1993; Yalden 1999). An 8th field in an anterior position can also occur in M1 ( Dienske 1969; Magalhães and Madureira 1980).
Marked geographical variation is found in size and color patterns of M. agrestis . M. agrestis from northern Europe ( Scotland, Scandinavia, northern Russia), the Alps, and Camargue ( France) are larger than those from central Europe (Krapp and Niethammer 1982). In England (Alibhai and Gipps 1991) and Sweden (Hansson and Jaarola 1989) size increases from south to north. Size also varies with elevation; larger M. agrestis are present at higher elevations in the area that was formerly Czechoslovakia (Krapp and Niethammer 1982) and in France and Portugal ( Almaça 1993). Individuals with darker dorsal pelage occur in Scandinavia and Scotland, and individuals with lighter pelage are found in central Europe, southern England, and northernmost Scandinavia (Corbet and Southern 1977; Krapp and Niethammer 1982).
Microtus agrestis males average slightly larger than females in external and cranial measurements in some populations ( Reichstein 1959; Krapp and Niethammer 1982), but exhibit little or no secondary sexual variation in size in other populations ( Wasilewski 1956; Gebczynska 1964; Saint-Girons 1973; Alibhai and Gipps 1991). Selected ranges of external and cranial measurements (mm; sexes combined) for adult M. agrestis from England (E, n = 38–46), France (F, n = 51–93), the Netherlands (N, n = 58), Switzerland (S, n = 17), and Portugal (P, n = 3–33) were: length of body, 115.0–121.0 (E), 85.0–130.0 (F), 78.0– 123.0 (S); length of tail, 25.0–46.0 (F), 29.5–46.0 (S); length of hind foot, 17.0–17.4 (E), 15.0–21.0 (F), 17.0–20.0 (S); length of ear, 9.0–14.0 (F), 10.0–13.0 (S); condylobasal length, 25.2–26.1 (E), 22.8–27.9 (F), 24.1–27.4 (S), 24.7–25.9 (P); zygomatic breadth, 12.1–15.7 (F), 12.1–16.1 (S), 12.6–14.1 (P); width of interorbital constriction, 3.1–3.9 (F), 3.3–3.8 (S), 2.3–3.2 (P); length of nasals, 5.5–7.8 (S), 5.9–7.0 (P); palatal length, 10.6– 15.0 (N); length of maxillary toothrow, 5.3–6.9 (F), 4.5–6.9 (N), 5.4–6.7 (S), 5.0–6.3 (P); length of mandibular toothrow, 5.2–6.7 (F), 5.4–6.6 (S), 5.0–5.8 (P); length of mandible, 12.4–17.0 (N), 14.0–17.3 (S), 13.6–15.9 (P—Dienske 1969; Saint-Girons 1973; Magalhães and Madureira 1980; Krapp and Niethammer 1982; Alibhai and Gipps 1991).
Ranges of cranial measurements (mm; sexes combined) of 26 M. a. levernedii (l) from Switzerland and France ( Madureira 1983), 13 M. a. agrestis (a) from Norway and Sweden ( Miller 1912), and 3 M. a. neglectus (n) from Scotland ( Miller 1912) were: condylobasal length, 26.6–28.4 (a), 25.0–25.4 (n); zygomatic breadth, 15.2–16.8 (a), 14.4–14.4 (n); width of interorbital constriction, 2.6–3.3 (l), 3.4–4.0 (a), 3.2–3.4 (n); width of postorbital constriction, 3.3–3.7 (l); nasal length, 7.2–8.0 (a), 6.8– 7.0 (n); palatal width, 1.5–1.8 (l); length of maxillary toothrow, 6.4–7.0 (a), 6.4–6.6 (n); length of mandibular toothrow, 6.2–6.8 (a), 6.2–6.4 (n); length of mandible, 14.9–17.6 (l), 16.8–18.0 (a), 15.4–16.2.(n). Cranial measurements (mm) of the type specimens of M. a. levernedii from Gard, France (female), and M. a. nigra (or niger) from Berne ( Geneva), Switzerland (male), respectively, were: condylobasal length, 28.0, 26.0; zygomatic breadth, 16.0, 14.8; width of interorbital constriction, 3.6, 3.4; nasal length, 8.0, 7.2; length of maxillary toothrow, 7.2, 6.4; length of mandibular toothrow, 7.0, 6.4; mandible length, 18.0, 16.6 ( Miller 1912). Ranges of body and cranial measurements from specimens collected in Europe were: length of head and body, 78–135 mm; length of tail, 18–49 mm (ca. 30% head-body length); length of hind foot, 16–20.5 mm, body mass 14–50 g (birth mass ca. 2 g); condylobasal length, 23–29 mm (sample sizes not given—MacDonald and Barrett 1993). Ranges of body measurements for individuals collected in Spain were: head and body length, 95–123 mm; length of tail, 25–44 mm; length of hind foot, 16–21 mm, mass 21–41 g (sample sizes not given— Gosalbez and Luque-Larena 2002).
Mean measurements (mm ± SD) for footprints of 25 M. agrestis were: forefoot print, 7.0 ± 0.7 in width and 7.6 ± 0.5 in length; hind foot print, 7.0 ± 1.0 in width and 9.1 ± 0.8 in length ( van Apeldoorn et al. 1993). Baculae (os penis) measurements (mm) of 2 adult males were: corpus length, 3.5, 3.2; basal width, 0.5, 0.4 ( Didier 1954).
DISTRIBUTION
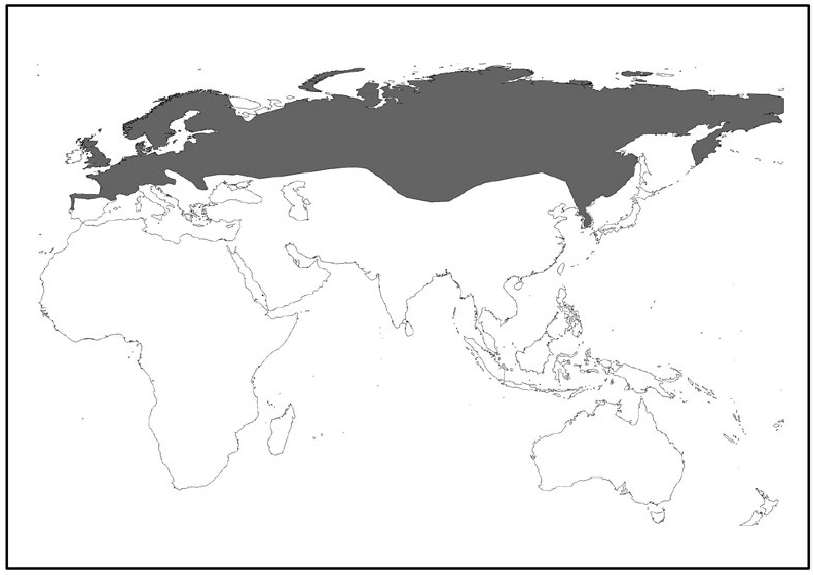
Distribution of Microtus agrestis ( Fig. 3 View Fig ) in northern and central Europe extends from Scandinavia to the Alps, Pyrenees, Galicia, and northern Portugal and from England, Scotland, and nearby small islands eastward to former Yugoslavia, southern Urals, Altai Mountains ( Mongolia), northwest China, and Lake Baikal ( Russia) regions ( Miller 1912; Ognev 1944; Corbet 1978; Krapp and Niethammer 1982; Brunet-Lecomte 1991; Gromov and Plyakov 1992; Mitchell-Jones et al. 1999; Musser and Carleton 2005). The species is absent from Iceland and Ireland ( Kryštufek et al. 2008).
The morphotype M. a. exsul comprises over 50% of specimens taken from the northern area of Russia, Scandinavia, and Scotland. In central Europe, the M. a. exsul type decreases to about 5%, and in southern Europe and the Alps, it never exceeds 20% of specimens collected (Krapp and Niethammer 1982).
FOSSIL RECORD
The 1st known fossil remains of Microtus agrestis are from the Upper Pleistocene of Europe (Kurtén 1968). Earlier records are mostly presented as the “ arvalis-agrestis ” group because these 2 species are only differentiated by M2, missing in most of the materials found. In the British Isles, M. agrestis is present from the Cromerian interglacial and Wolstonian period (penultimate glaciation) but never reached Ireland (Alibhai and Gipps 1991; Yalden 1999). Subfossil teeth collected from the island of Jura, Inner Hebrides, between 1,500 and 2,500 years ago, show a progressive simplification of the M1 pattern since that time ( Corbet 1975). The M. agrestis group (including M. agrestoides ) was present in the Middle Terrace stage, then absent until the Iglitham stage of the Late Pleistocene ( Hinton 1926).
Two geochronological forms from France were M. agrestis jansoni with larger size measurements of M 1 in the Middle Pleistocene and M. agrestis aubinensis with smaller size measurement values of M 1 in the Upper Pleistocene ( Chaline 1972). In Portugal, M. agrestis is found in Upper Pleistocene deposits of Caldeirão Cave in the center of the country ( Póvoas et al. 1992). In southern Poland, fossil remains of M. agrestis indicate that the species was present during the entire late Quaternary, including the Last Glacial Maximum ( Nadachowski 1984, 1989, 2001). Other fossils of M. agrestis are reported from Germany, Austria, Hungary, and Bulgaria ( Chaline 1972, 1974; Krapp and Niethammer 1982; Nadachowski 1984; Kolfschoten 1994), Hungary ( Jánossy 1986), southern Italy (Capasso-Barbato and Gliozzi 2001), and Eastern Europe including the Ural Mountains ( Markova et al. 1995). The presence of M. agrestis and other small mammals in sediments of the Middle Pleistocene from the Netherlands indicates interglacial conditions and a mainly wooded environment ( Kolfschoten 1991). Similar conclusions are drawn by Lopez-Garcia et al. (2011) for Burgos ( Spain). During the earlier stages of the last glacial period, M. agrestis was present in Asia ( Markova et al. 1995).
FORM AND FUNCTION
Form.— Microtus agrestis agrestis is one of the largest forms and M. a. gregarius is one of the smallest forms of M. agrestis . Brain case of M. a. levernedii is longer than that of M. a. agrestis ( Miller 1912) . Dental formula is: i 1/1, c 0/0, p 0/0, m 3/3, total 16. Specimens lacking the postero-internal M2 loop have been found from some Danish islands (Reichstein and Reise 1965). Differences in the frequencies of 25 epigenetic skeletal variants, involving skull, vertebrae, girdles, and long bones, were found among populations of M. agrestis from mid-Wales and Oxford, England (Berry and Searle 1963).
Feces of M. agrestis are small, green in color (Lawrence and Brown 1974), and 2.5 mm long (Alibhai and Gipps 1991). Olfaction is an important sense in communication. Caudal, preputial, and flank glands produce odors essential in intragroup and intraspecific recognition ( de Jonge 1980; Stoddart 1982).
Microtus agrestis has, in general, 2 annual molts, resulting in a sparse coat with coarse hairs in summer and a dense coat with fine hairs in winter. Thus, there is a change both in the number of hairs and in the amount of hair secreted by the follicles across seasons (Al-Khateeb and Johnson 1971). The molt appears to be adjusted to the environment by way of the endocrine system, with adrenal and thyroid hormones, as well as sex hormones, involved in the regulation ( Johnson 2008). Specimens born in southern England during the summer undergo a succession of molts before acquiring an adult coat; no hair growth occurs during December and January (Alibhai and Gipps 1991). Juveniles are darker gray than adults (Krapp and Niethammer 1982). Coat color does not vary throughout the year, although the summer coat is sparser with coarser guard hairs and fewer fine hairs than the dense winter pelage (Wettstein-Westersheimb 1959; Krapp and Niethammer 1982). M. a. macgillivrayii has a darker graybrown ventral pelage than central European or southern England forms (Krapp and Niethammer 1982; Alibhai and Gipps 1991). Reported mutant variations in coat color include agouti, pale, piebald, and melanism, though occurrence of melanism is very rare (Corbet and Southern 1977).
Function. —In field specimens, growth strategies are correlated with ambient temperatures. Growth is slow or stops altogether during winter, whereas body mass can increase by 50% during summer. In captivity, animals grew continuously throughout the year, both skeletally and in mass ( Bäumler 1979). Within wild populations variation of body size was suggested to be related to cycles of density ( Burthe et al. 2010). Animals with a body mass of 20 g consumed about 63.7 kJ of energy daily at 10°C ( Ferns 1976). Over the year, digestibility ranged between 33.6% and 67.8%, the highest values occurring in spring and summer ( Phillipson et al. 1983). Król and Speakman (2007) reported that Microtus agrestis responds to changes in photoperiod by altering digestive efficiency which is translated into changes in body mass and adiposity. M. agrestis may provide an attractive novel animal model for investigation of the regulation of body mass and energy homeostasis at both the organism and molecular levels (Król et al. 2005).
For M. agrestis captured in southern Sweden in late summer, average daily metabolic rate (ADMR) measured at 20°C was 3.95 ml O 2 g−1 h−1 (Hansson and Grodzinski 1970). Both ADMR and resting metabolic rate (RMR) depended on body mass ( W); at 20°C, ADMR = 19.54 W −0.53 and RMR = 34.7 W −0.63. ADMR of individuals housed in separate groups of either 4 or 8 animals, declined by 5–19% ( Hansson 1984). For M. agrestis maintained on a 16 h dark and 8 h light photoperiod, oxygen consumption peaked at the onset of darkness. RMR varied with ambient temperature ( T); Y = 9.42 − 0.21 X (Hansson and Grodzinski 1970).
Daily energy expenditure, determined by means of doubly labeled water, for free-living M. agrestis ranges from 1.8 to 4.5 times basal metabolic rate, with an average of 2.9 times basal metabolic rate ( Meerlo et al. 1997). Voles are apparently flexible in their thermogenic capacities allowing them to better survive under low ambient temperatures ( Jackson et al. 2001). McDevitt and Speakman (1994) reported that body mass, brown adipose tissue mass, white adipose tissue mass, and basal metabolic rate were found to be positively correlated with the length of time M. agrestis individuals were exposed to cold temperatures. Summer acclimatization is manifested as a significant decrease in basal metabolic rate, combined with a significant increase in body mass (McDevitt and Speakman 1996).
Short-term (ultradian) activity rhythm, between 2 and 4 h, was identified as associated with latitude or population dynamics. The ultradian rhythm is essentially a feeding rhythm and the periodicity is closely linked to food quality and energy needs ( Madison 1985), and perhaps avoidance of nest fouling (Alibhai and Gipps 1991).
Huddling by M. agrestis significantly reduces exposed surface area and increases heating. At an environmental temperature of 0°C, air temperature in metabolic chambers containing huddling groups was 11.8°C compared with 5.8°C when individuals were separated by partitions. Oxygen consumption, measured by open-circuit respirometry, of huddling groups was 23% lower than for animals separated by partitions within metabolic chambers ( Hayes et al. 1992). A 5°C increase in ambient temperature caused by huddling accounts for 55% of the energetic benefits for huddling M. agrestis ( Hayes et al. 1992) .
Vocalizations of M. agrestis include a faint, low-pitched mostly polysyllabic call and a range of chattering sounds ( Dienske 1969; Konig 1973). Sensitivity to sound shows a low-frequency peak at 9–10 kHz and a high frequency peak at 50 kHz for M. agrestis and other genera ( Apodemus , Clethrionomys , Gerbillus , Meriones , Mus , Peromyscus ). The high-frequency peak corresponds to frequencies of ultrasonic vocalization produced by the species; infant voles vocalize at 20–55 kHz ( Brown 1973). Ultrasonic calls of M. agrestis have fundamental frequencies of 35–50 kHz and a bandwidth of 5–10 kHz.
ONTOGENY AND REPRODUCTION
Ontogeny. —Young Microtus agrestis are naked and blind at birth and weigh about 2 g. Hair 1st appears between days 4 and 6 of life. Eyes open about the 10th day (Krapp and Niethammer 1982). Weaning occurs at 14–18 days, when young have attained an average weight of 8.9 g; young leave the nest at 21 days, and reach sexual maturity at about 6 weeks, depending on photoperiod and social factors ( Myllymäki 1977b; Spears and Clarke 1987; Ernest 2003).
Nestling mortality ranges from 20% in the spring to 50% during the “nutritional crisis” associated with limitations in food supply, when maturation and pregnancy rates also decline. Individual growth patterns, especially in females, and attainment of sexual maturity are closely correlated ( Erlinge et al. 1990; Alibhai and Gipps 1991). Maturation rate of females is negatively correlated with density of reproductive females (Pusenius and Viitala 1993a). Ergon et al. (2001) stated that seasonal decline in maturation rates is governed by changes in photoperiod.
Reproduction. —Breeding in Microtus agrestis is seasonal and extends from February to October in central Europe (Alibhai and Gipps 1991) and until November or December in other regions (Saint-Girons 1973; Hansson 1984; Erlinge et al. 1990) with a reproductive peak, June–August (Krapp and Niethammer 1982). This breeding pattern leads to recruitment of most young in summer or by early autumn. Breeding early in the year most likely occurs in those in better condition ( Beldomenico et al. 2008).
Testis development in M. agrestis raised under long photoperiods is rapid, and full sexual maturity is reached by 6–7 weeks. After this time the testis increased in weight to a plateau at about 70 days. Testis development under short photoperiods is much slower and when maturity is reached, for the majority of males between 4 and 6 months of age, testis weight was not as great as in animals of the same age kept in long photoperiods ( Grocock 1979). Males are considered fully sexually active when the testis have a cross-sectional area of> 40 mm 2 ( Andera 1981). Structural changes in male gonads that occur throughout the breeding season are related to the presence of luteinizing hormone (LH). Individuals from populations in England, exhibited increases in pituitary LH, testicular mass, and seminal vesicle mass between January and April. These values peaked from May to July, but declined from August to December ( Charlton et al. 1983). Body mass of males exhibited a significant positive correlation with reproductive success and dominance ( Nelson 1994; Nelson 1995a).
In spring, when females and males are densely aggregated, estrous cycles are asynchronous. When breeding females aggregate again in late summer, estrous cycles become more synchronous and males become territorial (Viitala and Pusenius 1990).
Ovulation is induced by mating or by the presence of sexually mature males through tactile, auditory, and visual stimuli ( Milligan 1974). Strange males can block the pregnancy of females previously mated with a stud male (Alibhai and Gipps 1991). Matrilineal kin clusters influence reproductive success ( Pusenius et al. 1998).
Experiments with M. agrestis under laboratory conditions demonstrated that selection for gonadal response to short photoperiods causes changes in growth rates and gonadal response after a single generation (Spears and Clarke 1988). Enclosure experiments and DNA fingerprinting revealed multiple paternity in 4 of 12 litters ( Berteaux et al. 1999).
Gestation is 18–21 days ( Corbet 1966). Average litter sizes at birth are 5–6 in Germany and 5–7 in Finland (sample sizes not given). In Poland, litter sizes at birth were: 2–7 ( n = 332, mean 4.25; for animals housed in a laboratory n = 32, mean 4.7). In former Czechoslovakia, range of litter size was 4.14–4.28 ( n = 292, corresponding to the total annual mean litter sizes of 3 different populations—Andera 1980; Krapp and Niethammer 1982). In England, litter size ranged from 2 to 7 in the wild and 4 to 6 in the laboratory, sample sizes not given (Alibhai and Gipps 1991). M. agrestis may have> 1 litter per year, average 3.6; inter-litter interval around 20 days (Alibhai and Gipps 1991; Corbet and Harris 1991; Ernest 2003).
Mean litter size based on embryo counts increases somewhat proportionately from small to larger females and is slightly higher than the number of young in litters ( Andera 1981). Number of litters produced, an index of reproductive success, varies with age and food availability ( Nelson et al. 1991). In Finland, litter size increases from south to north depending on phase of cycle, although litter size is one of the least variable parameters in the microtine cycle (Alibhai and Gipps 1991). Embryonic resorptions occur during the entire breeding season but rates are higher toward the end of the season ( Myllymäki 1977b).
Sex ratio of M. agrestis at birth is 1:1, although it becomes heavily weighted to females during the breeding season. This is especially true at peak abundance, due to differential dispersal of the sexes ( Myllymäki 1977b). In the laboratory under uniform conditions, sex ratios do not change ( Hansson 1987). Fecundity monitoring in M. agrestis was suggested as a useful biolomarker of environmental disturbance ( Caplen et al. 2004).
ECOLOGY
Population characteristics. —Maximum longevity of Microtus agrestis varies from 16 to 18 months in nature ( Myllymäki 1977b; Andera 1981) but is nearly 40 months in captivity ( Gebczynska 1964). Mortality is very high during the 1st month of life, especially during summer ( Myllymäki 1977b; Baumler 1979). Frequently, the proportion of adult or subadult females in populations is higher than males due to higher mortality accompanying male dispersal ( Myllymäki 1977a).
Reproductive traits of M. agrestis in southern Sweden’s Stockholm Baltic Archipelago island were compared to that of mainland voles all maintained in the laboratory over 3 years. Insular colonies, compared to mainland voles, were characterized by larger adult body size, more rapid growth, larger and heavier litters, and larger overall reproductive effort relative to body size. Both mainland and insular colonies are similar in gestation length, litter mortality, and age at sexual maturity. This suggests that larger body size and reproductive traits are coevolved traits of island colonization ( Ebenhard 1990).
Direct density-dependent regulation occurs in populations of M. agrestis (Klemola and Koivula 2002) . Cyclic and annual changes in population numbers relate to changes in body mass, survival, growth, and reproduction ( Myllymäki 1977a). Hematological parameters characterizing body condition of M. agrestis are affected adversely by high population densities ( Beldomenico et al. 2008). Genetic and behavioral mechanisms are insufficient alone to generate cycles; an additional, more relevant role has been attributed to habitat structure and dispersal movements (mediated by social behavior) between habitat patches of different quality ( Hansson 1977; Stenseth 1977) and climate changes ( Bierman et al. 2006). Dalkvist et al. (2011) concluded that habitat fragmentation and predator assembly jointly determine cycle length and amplitude in M. agrestis . However, lack of winter food supply is apparently a more direct factor in regulating density of populations than predation ( Huitu et al. 2003). Massey et al. (2008) reported that silica-based defenses in grasses may play a relevant role in driving vole population cycles; high silica levels may reduce vole body mass by 0.5% a day. Changes in reproduction do not drive cyclic fluctuations in M. agrestis populations but regulation of cycle populations can be disrupted by the influence of another grazer in the same trophic level ( Villar et al. 2014). Low survival of young has a role in the shift from the peak to the decline phase in late summer and early autumn (Norrdahl and Korpimäki 2002). Furthermore, differences in social behavior of juveniles born in spring-early summer or autumn may account for differences in the behavior of mature males and ultimately have an important effect on the annual population cycle ( Wilson 2008).
High population densities may be attained every 2–6 years ( Myllymäki 1970, 1977b; Krapp and Niethammer 1982). Peaks of abundance of M. agrestis in central Scandinavia occur at 3- to 4-year intervals. In Czech Republic, Gouveia et al. (2015) reported that M. agrestis dynamics are highly variable, although peaks of abundance occur regularly at intervals of 4–5 years, as elsewhere. Survival schedules, growth patterns, reproduction rates, population structures, and habitat dynamics are related to seasonal and cyclic phases ( Myllymäki 1977b). When population densities increase, damage occurs to natural vegetation and agricultural crops. The most common type of damage is girdling trunks of cultivated trees, usually under the snow cover. In horticulture, the apple tree is the commonest subject of injury, while, in forestry, several deciduous and conifer species are attacked ( Myllymäki 1977c).
Relative to southern noncyclic populations, cyclic populations in northern Sweden are more polygynous, have higher recruitment of breeding females, contain a higher percentage of females in litters, but show no female maternal effects. Differences between northern and southern populations may include traits such as chromosomal structure (Fredga and Hansen-Melander 1970), isozyme composition ( Rasmuson et al. 1977), dental patterns ( Reinwaldt 1968), genetically determined aspects of locomotor activity, and sensitivity to social stimuli ( Rasmuson et al. 1977; Nygren 1980a, 1980b), body size and condition (Hansson and Jaarola 1989), litter size ( Stenseth et al. 1985), and winter reproduction ( Hansson 1984).
Space use. —One of the most abundant and widespread microtine voles, Microtus agrestis , is found at elevations up to about 2,100 m (Saint-Girons 1973; Krapp and Niethammer 1982; Kryštufek et al. 2008). It prefers wet areas in meadows, riverside habitats, and forests with dense herbaceous cover (Corbet and Southern 1977; Hansson 1977; Kryštufek et al. 2008). Marginal habitats supporting lower population densities include woodlands, hedgerows, dunes, and moorlands (Alibhai and Gipps 1991). In central Scandinavia, the most productive habitats are permanent grassland patches with a high proportion of preferred dicotyledonous food plants ( Myllymäki 1977b). In Finland, Savola et al. (2013) reported that abundance of 2 voles, the bank vole ( Myodes glareolus ) and M. agrestis , are dependent on the current forestry practices. Marchi et al. (2013) stated that landscape structure and land management are relevant factors in shaping gene flow and genetic diversity of M. agrestis populations in agricultural areas in Denmark.
Presence of M. agrestis is indicated by well-formed runways at ground level (also used by other small mammal species), feces, and small mounds of short-nibbled grass leaves or stems ( Corbet 1966; Alibhai and Gipps 1991). Underground tunnels connected to surface runways are constructed, but are rarely very extensive ( Corbet 1966). Nests constructed of dry grass are usually round and are usually under roots or protected by dense vegetation ( Corbet 1966; Alibhai and Gipps 1991).
Home ranges of adult M. agrestis in Finland are between 400 and 800 m 2 (Krapp and Niethammer 1982); home ranges of males are estimated to be 600 m 2 and females about 300 m 2 (Alibhai and Gipps 1991). Home ranges of males increase to 1,000 –1,500 m 2 during the breeding season, becoming sympatric with those of females (Krapp and Niethammer 1982). Home ranges of females in suboptimal habitats are larger and overlap less than those in more suitable habitats (Pusenius and Viitala 1993b). There is a negative correlation between home-range size and density. No significant correlation between home-range size and body mass was found. Male residency was transient and recruitment of female offspring influences male residency ( Loughran 2007). The introduction of immigrants into resident populations of M. agrestis disturbs the social organization by reducing home-range size of resident females and increasing mortality of males (Riotte-Lambert et al. 2012).
Diet. —Diet of Microtus agrestis includes leaves and stems of grasses, dicotyledons, and ferns ( Hansson 1971; Evans 1973; Ferns 1976) and seeds of the more succulent plant species in spring and summer ( Phillipson et al. 1983). M. agrestis prefers easily available small umbrella herbs and seed producers rather than, for instance, rosette herbs, dwarf shrubs, and tall umbrella herbs ( Hjaelten et al. 1996). Exceptionally, animal prey (e.g., dipteran larvae) are taken (Krapp and Niethammer 1982; Kryštufek et al. 2008). Locally, preference for plant species seems to be related to palatability and nutrient content of available plants on the ground ( Wheeler 2005). Food quality, namely the availability of dietary proteins, is the primary extrinsic factor determining individual well-being, thereby ultimately influencing population growth ( Agrell et al. 1992). Forbes et al. (2014b) following a field experiment confirmed that the nutritional quality of forage may be a source of interannual variation in the density and growth rates of populations by directly influencing the proportion of females and of those that were pregnant or lactating. The type of diet reported for M. agrestis suggests that it may be a promising indicator to estimate the hazards to wildlife of heavy metal (Zn, Cu, Mn, Cd, Pb, Cr) pollution. However, previous studies showed that the response of M. agrestis to pollution is small and erratic ( Beardsley et al. 1970).
Diseases and parasites.— Microtus agrestis is a reservoir species for various infectious diseases transmissible to humans and game animals and it has been studied as a wild model of ecological immunology for better understanding immunity to infectious diseases ( Campillo et al. 1994; Jackson et al. 2011; Turner et al. 2011). M. agrestis harbors blood parasites Trypanosoma microti , Babesia microti , Grahamella microti , Hepatozoon microti ( Baker et al. 1963; Molyneux 1969; Healing 1981; Cox 1987), nonidentified Bartonella species ( Karbowiak et al. 2005), and tissue parasites Frankelia microti , Sarcocystis putorii , Toxoplasma gondii ( Tadros 1968; Cox 1987; Alibhai and Gipps 1991).
Microtus agrestis has been identified as the most likely maintenance host of Mycobacterium microti , responsible for several human cases of M. microti tuberculosis not only in Great Britain but in other Western European countries ( the Netherlands, Germany, and France—Kipar et al. 2014). Broom and Coghlan (1958), Alibhai and Gipps (1991), Twigg (1980), and Telfer et al. (2010) summarized additional bacteria isolated from M. agrestis , including Leptospira (Pomona) , Yersinia pseudotuberculosis , Anaplasma phagocytophilum , and Corynebacterium kutscheri .
There is also evidence that Tula virus, lymphocyte choriomeningitis virus, Ljungan virus, and cowpox virus are transmitted by M. agrestis ( Scharninghausen et al. 2002; Burthe et al. 2008; Schmidt-Chanasit et al. 2010; Forbes et al. 2014c; Salisbury et al. 2014). Burthe et al. (2008) reported a negative correlation between cowpox virus infection and M. agrestis survival, both at the individual level and population level and highlight for the 1st time an association between an endemic microparasite and processes of demographic importance in a wild rodent population. Telfer et al. (2010) stated that the risk of infection of M. agrestis by a particular microparasite (viruses, bacteria, and protozoa) is highly influenced by previous infections.
Parasites in the intestinal tract of M. agrestis include the Protozoans Chilomastix , Eimeria falciformis , Entamoeba muris , Giardia muris , Hexamita muris , Spironucleus muris , Trichomonas microti , T. muris , and Cryptosporidium ( Cox 1987; Alibhai and Gipps 1991; Laakkonen et al. 1994, 1998). Cestodes reported are Anoplocephaloides dentata , Aprostatandrya macrocephala , Catenotaenia cricetorum , C. pusilla , Corrigia vitta , Cysticercus taeniaetaeniaeformis , Hydatigena taeniaeformis , Hymenolepis asymetrica , H. diminuta , Mesocestoides lineatus , Paranocephala dentata , P. omphalodes , P. gracilis , Rodentolepis ampla , Taenia laticollis , T. mustela , T. polyacantha , T. taeniaeformis , and T. tenuicollis ( Wiger 1977; Lewis 1987; Alibhai and Gipps 1991; Haukisalmi et al. 1994). Nematodes include Aspiculuris tetraptera , Capillaria murissylvatici , Heligmosomum costellatum , Heligmosomoides glareoli , H. laevis , H. polygyrus , Pelodera strongyloides , Protospirura muris , Syphacia nigeriana , S. obvelata , and Trichurus muris ( Wiger 1977; Lewis 1987; Alibhai and Gipps 1991; Haukisalmi et al. 1994).
The presence of intestinal nematode parasites Heligmosomidae apparently does not influence M. agrestis population growth ( Forbes et al. 2014a). However, when M. agrestis is infected with B. microti , Giardia microti , or T. microti during the first 2 weeks of pregnancy, resorption or abortion of fetuses or maternal death may occur. During the peak population density phase, parasitism is thought to contribute to the rapid decline of numbers ( Wiger 1977).
Ectoparasites on M.agrestis are fleas Amalaraeus penicilliger , Ctenophthalmus nobilis , Histrichopsylla talpae , Megabothris turbidus , M. walkeri , M. rectangulatus , Nosopsyllus fasciatus , Peromyscopsylla spectabilis , Rhadinopsylla integella , R. pentacantha , Typhloceras poppei , Haplopleura acanthopus , and ticks Ixodes ricinus , I. triunguliceps (Alibhai and Gipps 1991) . The incidence of I. ricinus ticks (larvae and nymphs) in M. agrestis in France peaks from March to October ( L’Hostis et al. 1996). I. ricinus and their hosts, namely M. agrestis , are responsible for maintaining tick-borne encephalitis endemic in central and northern Europe ( Tonteri et al. 2011). Brown et al. (2008) also pointed out the relative importance of I. ricinus and I. trianguliceps in the transmission of A. phagocytophilum and B. microti among M. agrestis populations in the United Kingdom, which serve as reservoir hosts for both pathogens.
Mites include Eulaelaps stabularis , Haemogamasus nidi , Laelaps hilaris , Listrophorus leuckarti , and Trombicula autumnalis (Alibhai and Gipps 1991) . Recorded fungi is Trichophyton persicolor ( English 1966; Alibhai and Gipps 1991); a known lung parasite is Pneumocystis carinii (Laakkonen and Sukura 1997) . English (1966) isolated T. persicolor , thought to be identical to human ringworm, from tail lesions of M. agrestis hirtus .
Interspecific interactions.— Microtus agrestis is the most important dietary part of more species of predators than perhaps any other mammal due mainly to its numerical abundance ( Corbet 1966). Avian predators of M. agrestis include barn owl ( Tyto alba ), common buzzard ( Buteo buteo ), golden eagle ( Aquila chrysaetos ), common kestrel ( Falco tinnunculus ), Northern harrier ( Circus cyaneus ), long-eared owl ( Asiootus ), Eurasia pygmy owl ( Glaucidium passerinum ), shorteared owl ( Asio flammeus ), tawny owl ( Strix aluco ), and Ural owl ( Strix uralensis — Corbet 1966; Andersson and Erlinge 1977; Madureira 1979; Alibhai and Gipps 1991; Korpimäki 1992; Mappes et al. 1993; Viitala et al. 1995; Dyczkowski and Yalden 1998; Koivula et al. 1999; Koivula and Korpimaki 2001; Redpath et al. 2002). In Finland, heavier male M. agrestis and lighter-weight subordinates are more vulnerable to predation by Ural owls than females ( Mappes et al. 1993; Haukisalmi et al. 1994; Korpimäki et al. 1996a). However, the probability of individual M. agrestis being predated was not related to infections by helminth species or overall parasite infection levels according to Haukisalmi et al. (1994). Kestrel predation has little impact on pregnancy rates, mean litter sizes, or growth rates ( Klemola et al. 1998). Predatory kestrels appear to be able to differentiate between M. agrestis and other voles and reproductive categories based on scent markings ( Koivula et al. 1999). In a field study, Petty et al. (2000) concluded that over a 23-year period the smallscale synchrony in M. agrestis abundance observed in northern England is unlikely to be caused by avian predators but instead by forestry practices.
Mammalian predators include European badger ( Meles meles ), least weasel ( Mustela nivalis ), European pine marten ( Martes martes ), European polecat ( Mustela putorius ), feral cat ( Felis catus ), red fox ( Vulpes vulpes ), ermine or stoat ( Mustela erminea ), and wildcat ( Felis silvestris — Corbet 1966; Andersson and Erlinge 1977; Alibhai and Gipps 1991; Dyczkowski and Yalden 1998). Adders ( Vipera berus ) prey on M. agrestis (Forsman and Lindell 1997) . Large populations of specialist predators markedly influence summer declines of cyclic populations of M. agrestis in northern Europe (Korpimäki and Norrdahl 1998).
Dyczkowski and Yalden (1998) estimate that total predator consumption of voles/year is 980,000, while productivity is estimated to be as high as 982,000/year. Two specialist vole predators, weasel and kestrel, and 2 generalist predators, red fox and feral cat, consume 85% of the total M. agrestis taken by predators. A recent study by Huitu et al. (2003) concluded that winter food shortage is a more direct factor in regulating density of M. agrestis populations than predation.
BEHAVIOR
Reproductive behavior. —The normal mating system of Microtus agrestis in central Finland (and probably elsewhere) is polygyny with strictly territorial males. Intrasexual competition and spacing behavior occurs among males in response to density and positioning of females ( Nelson 1995b). M. agrestis reduces competition with other males by marking copulatory domans; thus, male density influences spacing behavior of male M. agrestis ( Nelson 1995b) . In optimal habitats at high density, promiscuous multi-male groups occur (Viitala and Pusenius 1990; Pusenius and Viitala 1993a).
Comparing nest construction by males and females, Redman et al. (1999) concluded that nests built by males had greater insulation and thicker walls than those built by females, thus allowing better protection from ambient temperatures. Care of young M. agrestis is by mothers only (MacDonald and Barrett 1993). Maternal investment is clearly related to the social system; the sex with the largest number and highest quality of young was that which was not limited in number by territoriality. However, investment also depends on time until maturation of young. Young females that are born early in the summer mature and breed in their year of birth, although this varies greatly. Late-summer young do not mature until the following year ( Nelson et al. 1991; Hansson 1992).
Sex ratios between generations may be regulated by several factors such as maternal condition and the prevailing social system ( Hansson 1987; Nunney 1993). Cyclic populations of M. agrestis seem more polygynous than noncyclic ones and more females in litters had been reported from cyclic than from noncyclic populations ( Hansson 1987, 1992). Deviations in sex ratios have been justified with the general idea that females invest most heavily in the sex with the highest reproductive success (largest number of young and highest quality—Hans-son 1987).
In a lab experiment, Clarke (1953) reported that resident M. agrestis attack strange conspecifics. As a result of these attacks there was a pronounced increase in the weight of the adrenal glands and the spleen, and a decrease in the weight of the thymus of the strange voles, justified by a disturbance of the adrenopituitary system and a decrease of the body condition of voles.
Communication. —In Microtus agrestis , shorter (67 ms), higher frequency calls have been detected during attack while a few longer (100–125 ms), lower frequency calls were detected during approach ( Kapusta et al. 2007). In adults, the emission of ultrasonic pulses seems to be modified by sexual and parental experience of M. agrestis in heterosexual encounters; the environment in which the animals meet also seems to affect ultrasound emission (Mandelli and Sales 2004).
Neonatal M. agrestis usually emit the tonal calls that are typical of infant distress vocalizations, but they also make soft clicking sounds during the 1st few days of life (Mandelli and Sales 2004). Infant M. agrestis vocalize more than bank vole and the water vole ( Arvicola terrestris ). Mid-aged M. agrestis maximal vocalization occurs at 5°C with mean sound pressure level of 890 dB and mean of 1,291 vocalizations/20 min. The risk of hypothermia leads to a vocalization response in infant voles ( Blake 1993). Most infant voles vocalize ultrasonically when isolated. Calls reflect changes in physical development of infants and in their external environment ( Blake 1993; Mandelli and Sales 2004).
Miscellaneous behavior. —Activity of Microtus agrestis is generally nocturnal in summer, probably the result of higher daytime temperatures, and more diurnal in winter due to colder nights. Juveniles tend to be more nocturnal than adults ( Bäumler 1975). General activity levels decrease with rainfall associated with low temperatures ( Baumler 1975; Lehmann 1976; Alibhai and Gipps 1991), but M. agrestis does not hibernate and is active year-round ( Nowak 1999).
During peak density, diurnality prevails in M. agrestis ; distinct activity maxima occurred at sunrise and sunset and a short-term rhythm of activity (rest and foraging) of 2–2.5 h was discernible (Halle and Lehmann 1992; Nowak 1999). In late population declines, a phase-shift from diurnality to nocturnality occurs in July–August. During low density and early increase phases, diurnality prevails again, although daily activity patterns are not as regular as during the peak phase (Halle and Lehmann 1992). Experimentally increasing density to evaluate spacing behavior of male M. agrestis resulted in smaller, more overlapping home ranges, less movement, less activity, and increased competition among males with seasonal variation ( Agrell et al. 1996). Density of females influences spacing behavior of males. At high female density, males have smaller home ranges and move shorter distances ( Nelson 1995a; Agrell et al. 1996). In several populations, a decline in density occurs in mid-summer ( Myllymäki 1977b). This period is characterized by increasing territoriality, decreasing juvenile survival, and decreasing population density (Viitala and Pusenius 1990).
During the nonbreeding period at higher densities, both males and females move shorter distances than during the breeding season at lower densities. Distances moved do not differ between nonbreeding and breeding periods in increasing and declining population densities ( Sandell et al. 1991). Juveniles rarely attain sexual maturity while resident in their natal area; 8% of 106 males ( 9 males) and 22% of 134 females ( 29 females) attained sexual maturity within their natal home ranges, whereas the remaining juveniles dispersed before sexual maturity ( Sandell et al. 1990). Among individuals re-trapped as adults, 58% of males and 23% of females dispersed; distances moved were> 1 home-range diameter. Mean distance moved by males ( 58.5 m) exceeded that of females ( 28.6 m). Distances moved by males were negatively associated with total density and density of adult females, but not male density. Female movements were not correlated to population density. No relationship between sex ratio of the population and distance moved were observed for either sex ( Sandell et al. 1990).
Microtus agrestis was found to modify spatial behavior, depending on season, as a reaction to predation risks by carnivore predators; the basic antipredatory reaction limits their locomotory activity (Borowski and Owadowska 2010). Distributions of distances moved for both sexes fits a normal geometrical distribution ( Sandell et al. 1990).
Female clans of mothers and daughters may occur (Viitala and Pusenius 1990). Young females disperse from maternal home range less often than do subadult males or breeding females ( Myllymäki 1977a; Viitala 1977).
Small mammalian carnivores have a stronger impact on M. agrestis prey population dynamics than avian predators and kill more females than males (Norrdahl and Korpimäki 1998). Small mustelids also selectively kill more females than males ( Klemola et al. 1997). Odor or scent marks left as part of reproductive activities increase risk of predation by mammalian predators more than UV visibility of scent marks (Koivula and Korpimäki 2001). Under simulated predation risk using mustelid odors, nonmating behavior of both female and male M. agrestis changed. Moreover, mass of both sexes decreased (Kostela and Ylönen 1995). Predation by small carnivores may result in M. agrestis increasing their survival by reducing mobility (traveling less—Norrdahl and Korpimäki 1998). Radiocollars do not markedly alter predation ( Korpimäki et al. 1996b).
GENETICS
Microtus agrestis has a diploid number (2n) of 50 chromosomes with a fundamental number (FN) of 54 (Zima and Král 1984). Only the largest autosome, a telo- or acrocentric, and the smallest autosome, a metacentric, are identifiable in unbanded preparations. The other 22 autosomes are acrocentrics and form a continuous series of decreasing lengths. The C- and G-band patterns of metaphase chromosomes (Cooper and Hsu 1972; Pera 1972) and the NOR-band patterns have been described (Goodpasture and Bloom 1975; Sperling et al. 1987; Ashley et al. 1989).
The sex chromosomes of M. agrestis are extremely large due to the accumulation of huge blocks of constitutive heterochromatin ( Matthey 1949; Wolf et al. 1965; Ohno 1967). The submetacentric X chromosome is almost 3 times longer than the largest autosome, and the acrocentric Y chromosome is as long as the long arm of the X chromosome. A variant metacentric X reaching 3.7 times the length of the largest chromosome was found in Portuguese populations ( Giménez et al. 2012). Molecular characterization of M. agrestis sex chromosomes is discussed in the context of pairing, evolution, and phylogeny ( Modi 1987, 1993; Nanda et al. 1988; Borodin et al. 1995; Kalscheuer et al. 1996; Nesterova et al. 1998).
A variant subtelocentric Y chromosome, known as the Lund Y chromosome, found in individuals collected from southwestern Sweden (Hansen-Melander 1964) results from a pericentric inversion (Fredga and Jaarola 1997). In M. agrestis from Portugal, the Y chromosome presents an intermediate morphology between the standard Y (“minute short arm”) and the Lund Y ( Giménez et al. 2012). X and Y chromosomes do not pair during meiosis in either of the 2 Y chromosome types ( Ashley et al. 1989). In an extensive geographic survey comprising 353 males from northern and central Europe, the distribution of the Lund Y chromosome was restricted to southwest Sweden (Fredga and Jaarola 1997).
Allozyme variation in 3 M. agrestis populations from Sweden contained a very high level of average heterozygosity (0.19–0.22) and an excess of homozygotes ( Nygren 1980b, 1980c; Nygren and Rasmuson 1980). One of these populations was reanalyzed with a more representative battery of enzymes, and only 3 of the 21 loci were polymorphic with no evidence of inbreeding ( Frykman 1988).
Restriction enzyme analysis of mitochondrial DNA in M. agrestis from northern Europe revealed distinct geographic patterns of diversity and lineage distribution (Jaarola and Tegelström 1995, 1996). Overall mtDNA diversity was high, but showed a pronounced geographic heterogeneity. Two major mtDNA lineages were identified as differing by 2% in nucleotide sequence. A wide contact zone between the 2 M. agrestis mtDNA groups was found in a suture zone in northern Sweden where several mammalian intraspecific zones of contact occur (Jaarola and Tegelström 1995; Jaarola et al. 1999).
Strong but complex phylogeographic substructuring was observed within both major mtDNA groups (Jaarola and Tegelström 1995, 1996). A population characterized by fixation of 2 unique sex-specific genetic markers, the Lund Y chromosome, and a specific mtDNA sublineage was identified in southwestern Sweden (Jaarola and Tegelström 1996). This Lund population showed the imprints of a population that underwent a bottleneck or founder event and probably originated about 12,000 years ago. A contact zone between the Lund population and a standard population with the 2 sex-specific markers was identified. The mtDNA and Y chromosome clines exhibited a rarely observed lack of coincidence, and as a consequence, only 1 of 2 possible male hybrid classes was observed ( Jaarola et al. 1997).
A survey of the mtDNA phylogeographic pattern across the species range, using sequence analysis of the whole cytochrome- b gene (1,140 bp), demonstrated that M. agrestis is divided into 3 major phylogenetic lineages that exhibit strict geographic subdivision (Jaarola and Searle 2002). The northern and central part of the species’ range is divided into a western and an eastern phylogeographic group. These 2 groups were identical to those previously identified in northern Europe by restriction site data (Jaarola and Tegelström 1995). The 3rd phylogeographic group in M. agrestis is present in southern Europe in the Iberian Peninsula, southernmost France, Switzerland, northernmost Italy, Slovenia, eastern Hungary, and Croatia (Jaarola and Searle 2002, 2004; Hellborg et al. 2005). Mitochondrial DNA clock estimates suggest that the western and eastern phylogeographic groups separated during the last glacial period, whereas the southern population dates back at least 500,000–900,000 years. The degree of differentiation of the southern mtDNA lineage (5–6%) is similar to that observed between sibling species in Microtus (Jaarola and Searle 2004; Jaarola et al. 2004). The southern population can also be detected with paternally (Y chromosome) and biparentally (X chromosome) inherited DNA sequences ( Hellborg et al. 2005). Together, the genetic data suggest that the morphologically cryptic but ancient and evolutionary distinct southern M. agrestis represents a separate species (Jaarola and Searle 2002, 2004; Hellborg et al. 2005).
Beysard et al. (2012) studied a contact zone between the southern and northern (i.e., eastern and western) M. agrestis lineages in the Swiss Jura Mountains using maternally, paternally, and biparentally inherited genetic markers. Despite a strong genetic structure in the contact zone, some hybrids and gene flow occurred. The extent of introgression varied with the mode of inheritance and was asymmetric, occurring only from the northern to the southern lineage. These results can be explained by neutral processes linked to differences in effective population sizes and sex-biased dispersal rates. However, it is possible that Haldane’s rule contributes to the pattern observed.
Herman and Searle (2011) showed that the eastern and western mtDNA lineages can be further divided into 6 lineages. Using new and published cytochrome- b gene sequences and an extremely fast molecular clock, they estimated that a single mitochondrial ancestor occurred around 24,000 years ago. The 6 regional lineages originated about 12,000 years ago from bottlenecked populations.
Based on a multilocus approach, Paupério et al. (2012) demonstrated a 3rd major lineage in M. agrestis in Portugal. Coalescent analyses supported 3 evolutionary units that may represent cryptic species: the northern and southern (as described previously) and Portuguese. Divergence among these units was estimated to have occurred during the last glacial period. Herman et al. (2014) calculated the rate of the mitochondrial cytochrome- b molecular clock using the postglacial colonization of Scandinavia by M. agrestis and a well-established cytochrome- b genealogy for the northern lineage.
CONSERVATION
Microtus agrestis is present in a large number of protected areas throughout its wide range. It is considered a species of “Least Concern” by the International Union for Conservation of Nature and Natural Resources and as such no specific conservation measures are required ( Kryštufek et al. 2008). Locally it can be rare in marginal parts of its range although in some other areas M. agrestis may cause considerable damage to grassland and fruit tree plantations, wood-stemmed ornamental trees, and forest trees. In arable land, M. agrestis cuts the plant stem close to the ground, then consumes the stem and leaves (Alibhai and Gipps 1991; Zima 1999).
Productivity estimates of M. agrestis in Britain were estimated to be between 677,000 and 982,000 per year depending on calculation method (Dyczkowski and Yalden 1998) though varying habitat availability has undoubtedly been a factor in overall vole population dynamics.
Charles (1981) reported that M. agrestis causes infrequent damage to British flora and is not usually regarded as an important economic pest of British coniferous forests. However, Kanervo and Myllymäki (1970) stated that M. agrestis has been an important pest in Great Britain and Germany. In Scandinavia, vole problems continue at 4-year cyclic intervals as M. agrestis reaches high population numbers. In Finland and Sweden, economic loss due to damage from M. agrestis to apple trees and horticultural plants during the 1950s and 1960s was minimally estimated at $2 million, each, over 13 years. Forestry damage in Finland and Norway during the 1960s was estimated at $120,000 each; estimates of damage in Sweden were much higher. Heavy losses to stored hay especially during the 1966–1967 winter were recorded in Sweden and Finland. Outbreaks were mostly synchronous during the 1950s and 1960s in Finland, Sweden, and Norway, though northern latitudes diverged from this pattern. Overall, damage resulting from M. agrestis is said to be the most serious pest problem to Finnish, Swedish, and Norwegian horticulture (Kanervo and Myllymäki 1970).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Microtus agrestis ( Linnaeus, 1761 )
| Mathias, Maria Da Luz, Hart, E. Blake, Ramalhinho, Maria Da Graca & Jaarola, Maarit 2017 |
Microtus agrestis enez-groezi
| Heim de Balsac and de Beaufort 1966: 638 |
Microtus agrestis armoricanus
| Heim de Balsac and de Beaufort 1966: 638 |
Microtus agrestis carinthiacus
| Kretzoi 1958: 57 |
Microtus agrestis argyropuli
| Ognev 1950 |
Microtus agrestis scaloni
| Heptner 1948: 710 |
Microtus agrestis argyropoli
| Ognev 1944: 179 |
Microtus agrestis argyropoli
| Ognev 1944 |
Microtus agrestis pallida
| Melander 1938: 74 |
Microtus agrestis ognevi
| Skalon 1935: 11 |
Microtus agrestis ognevi
| Skalon 1935 |
Microtus agrestis
| Wettsteini Ehik 1928: 197 |
Microtus agrestis estiae
| Reinwaldt 1927: 13 |
Microtus hirtus orioecus
| Cabrera 1924: 8 |
Microtus agrestis pannonicus
| Ehik 1924: 76 |
Microtus agrestis tridentinus
| Dal Piaz 1924: 10 |
Microtus agrestis punctus
| Montagu 1923: 868 |
Microtus agrestis fiona
| Montagu 1922: 940 |
Microtus agrestis mial
| Barrett-Hamilton and Hinton 1913: 364 |
Microtus agrestis luch
| Barrett-Hamilton and Hinton 1913: 366 |
Microtus agrestis macgillivrayii
| Barrett-Hamilton and Hinton 1913: 831 |
Microtus agrestis arcturus
| Thomas 1912: 398 |
Microtus agrestis mongol
| Thomas 1911: 759 |
Microtus agrestis insul
| Lydekker 1909: 74 |
Microtus agrestis exsul
| Miller 1908: 201 |
Arvicola agrestis angustifrons
| Fatio 1905: 191 |
Arvicola agrestis latifrons
| Fatio 1905: 194 |
Arvicola agrestis var. nigra
| Fatio 1869: 241 |
Arvicola rozianus
| Bocage 1865: 7 |
Arvicola levernedii
| Crespon 1844: 73 |
Lemmus insularis
| Nilsson 1844: 34 |
Lemmus insularis
| Nilsson 1844 |
Arvicola neglecta
| Jenyns 1841: 270 |
Arvicola hirta
| Bellamy 1839: 373 |
Mus arvalis nigricans
| Kerr 1792: 239 |
Mus gregarius
| Linnaeus 1766: 84 |
Mus agrestis
| Linnaeus 1761: 11 |
Mus agrestis
| Linnaeus 1761 |