
Goniopholis, AND
|
publication ID |
https://doi.org/10.1111/j.1096-3642.2011.00709.x |
|
persistent identifier |
https://treatment.plazi.org/id/03B40213-FFEC-FFAF-FF00-56D6FBC7FD3A |
|
treatment provided by |
Valdenar |
|
scientific name |
Goniopholis |
| status |
|
GONIOPHOLIS AND GONIOPHOLIDIDAE
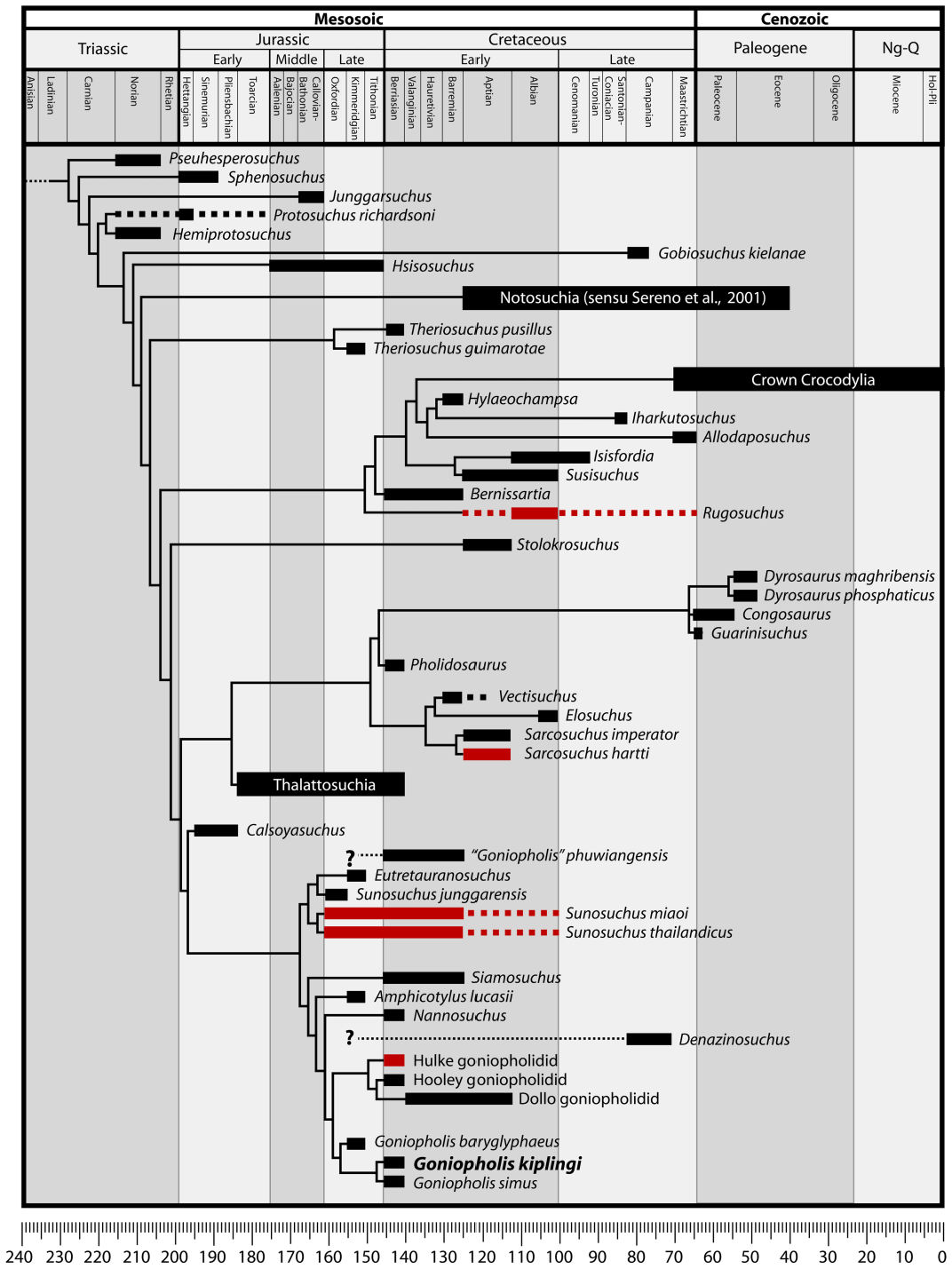
Worldwide reports of Goniopholis have been based mostly on teeth or fragmentary material, and a wide sample of teeth and osteoderms can be found in most palaeontological collections in Europe, North America, Brazil, and Asia. Indeed, the morphologies of teeth and dermal scutes contrast with the pattern seen in basal crocodylomorphs, notosuchians, eusuchians, and most thalattosuchians. However, in the case of tooth morphology, overall the same pattern (i.e. robust crowns, keeled, with intense well-defined ornamentation that becomes anastomozed near the apex, nonziphodont to false-ziphodont) can be found in many goniopholidids, pholidosaurids, and dyrosaurids. Among thalattosuchians, at least the teleosaurid Machimosaurus has remarkably similar tooth crown morphology. Dermal scutes, on the other hand, seem to allow taxonomic assignment within certain groups (D. Schwarz-Wings, pers. com. 2010), but isolated material still provides only limited clues for identification. Indeed, not much is known of the intraspecific (or intrageneric) variability of teeth and components of dermal armour, a problem that hinders comparison between taxa, and identification of truly meaningful characters. The use of tooth morphology is particularly important, because it is the sole element supporting the Goniopholis paulistanus (Upper Cretaceous, Brazil). The few cases where the crown morphology provides highly distinctive characters that allow specific/generic identification are usually restricted to notosuchian taxa with remarkable heterodonty (e.g., Candidodon , Chimaerasuchus ). Within neosuchia, perhaps the best exemple of autapomorphic teeth is Iharkutosuchus , but it must be noted that the taxon was described based on plentiful skull material (see Osi & Weishampel, 2009). It was also previously pointed out that tri-faceted laminar ziphodont crowns – so far – only occur in the metriorhynchid genus Geosaurus ( Young & Andrade, 2009; Andrade et al., 2010). The identification of tribodontic teeth from the Purbeck beds to Bernissartia is another example, but it was also regarded as extremely tentative (see Salisbury, 2002; and references within). Salisbury et al. (1999) have already recognized that the variability present in tooth morphology in goniopholidids prevents the use of such characters to support identification of species. Apart from the examples above, descriptions that state presence of ornamentation, rhomboid apex, or presence of ‘bicarinate teeth’ in most cases cannot consubstantiate discrete characters that can – based on crown morphology alone – secure a definitive specific/generic assignment, let alone the erection/maintenance of a new taxon. Still, crown morphology remains an overall useful tool for gross recognition of morphotypes (see Andrade et al., 2010), but any taxonomic assignment or species definition must also be supported by a phylogenetic analysis, or other meaningful associated remains. Based on the above, tooth morphology is seen here as neither sufficiently precise to provide a secure assignment to Goniopholis / Goniopholididae , nor to provide a diagnostic combination of features. Therefore, its usage alone is disregarded here. In the absence of meaningful information that allows the placement of this type of material (i.e., teeth and other isolated material) in a phylogenetic analysis, the material itself cannot be used to support taxonomic assignment. In these cases, the phylogenetic analysis cannot effectively provide clues to the possible relationships, simply because of scarceness of details. The removal of these specimens from the genus is important, for they offer feeble support for the wide geographical and stratigraphical distribution of Goniopholis (previously global, ranging from Upper Jurassic to Upper Cretaceous).
Trustworthy accounts of Goniopholis are limited to Laurasian sediments (see a reappraisal of brief Gondwanan reports below). Considering the new definition for the genus presented herein, the genus itself is limited to Europe, and ranges from the Kimmeridgian ( G. baryglyphaeus ) to the Berriasian ( G. simus , G. kiplingi ). The presence of Goniopholis in Upper Cretaceous sediments was mostly based on Denazinosuchus (Kirtland Formation, USA), a relation that is rejected by our analysis. The putative occurrence of Goniopholis in South-East Asia ( Thailand) was based on Su. phuwiangensis , and is also rejected.
Two cases of co-occurrence of ‘broad’ and ‘narrowsnouted’ goniopholidids in the same formation are now known. The first case is Amphicotylus and Eutretauranosuchus , both from the Morrison Formation (Morrison Basin), Kimmeridgian of the USA. The second case becomes evident with the recognition that Su. phuwiangensis is a ‘ Sunosuchus- like’ taxon, and comes from the Sao Khua Formation of Thailand ( Khorat Group), pre-Aptian of Thailand. Although further research is needed to explore the stratigraphical distribution and spatial range of these taxa, both cases may indicate niche partitioning in the group.
Overall, goniopholidids represent an important radiation from the Early Jurassic ( Calsoyasuchus ) to the Late Cretaceous ( Denazinosuchus ) of the USA (~126 Mya), which has a particularly diverse record through the Late Jurassic- Early Cretaceous boundary ( Fig. 14 View Figure 14 ), spreading through Asia and Europe. The clade of European goniopholidids remains a poorly understood group, but is here recovered as monophyletic, with Dollo’s specimen indicating presence up to late pre-Aptian times. Interestingly, Sunosuchus has no record in Europe, and the genus is limited to Asia. The earliest record of Sunosuchus is from the Middle Jurassic of China ( Su. shunensis ), and the latest are reports from the?late Early Cretaceous of Mongolia/ Russia (see Schellhorn et al., 2009 for a detailed review). In general, records of Sunosuchus -related taxa are older than the records from Goniopholis - related taxa. However, the genus Sunosuchus remains poorly explored and demands appropriate revision, which may reveal as much diversity as seen in Goniopholis sensu lato.
The Gondwanan record of Goniopholis and Goniopholididae
Gondwanan reports of Goniopholis are much rarer than Laurasian ones. However, a couple of reports originate from Brazil. ‘ Goniopholis ’ hartti ( Marsh, 1869) from the Lower Cretaceous is in fact a member of the genus Sarcosuchus . The results herein confirm for the first time, on the basis of a phylogenetic analysis, the taxonomic amendment proposed by Buffetaut & Taquet (1979). Sarcosuchus hartti is either the sister group of Sa. imperator , or the first is a senior synonym of the second, and not part of the genus Goniopholis . The second report is G. paulistanus Roxo 1936 , from the Upper Cretaceous Bauru Group ( São Paulo State, Brazil). The material is limited to two tooth crowns and a fragment of a (right) tibia, and lacks diagnostic characters that can provide a reliable link to the genus Goniopholis , or even to the family Goniopholididae (see ‘Remarks on diagnosis’ of Goniopholis ), regardless of its taxonomic status. These specimens of G. paulistanus are here treated as Neosuchia incertae sedis.
These Gondwanan accounts for Goniopholis (either sensu lato or stricto) are therefore based on information that cannot be truly linked to this genus. By solving these problematic reports, it is confirmed that Goniopholis is not known from Gondwana. Furthermore, the Bauru specimens assigned to the genus erroneously indicated the presence of the genus in the Upper Cretaceous of South America. Although possible, it seems unlikely that Goniopholis existed in the post-Barremian of Gondwana, and so far no evidence can corroborate this idea. Furthermore, there is plenty of evidence for the presence of tethysuchian taxa in Gondwana (e.g. Sarcosuchus , Meridiosaurus , Elosuchus , Dyrosaurus , Guarinisuchus ), and their presence can arguably be used to explain at least some of the Gondwanan accounts of ‘ Goniopholis ’, based on fragmentary material. However, further research is needed to understand fully the palaeobiogeographical distribution of these clades.
The family Goniopholididae may have been present in Gondwana, although most notes refer to old reports on taxa with similar tooth morphology. This is the case for Itasuchus jesuinoi Price, 1955 , which was originally included in the family but is more often considered as a trematochampsid (e.g. Buffetaut, 1991). At least in South America, there is no further reference to the family Goniopholididae (see Candeiro & Martinelli, 2006). However, goniopholidids may have been present in Gondwanan territory, as preliminarily reported by Sereno (2009). The material corresponds to an undescribed, fairly complete ‘ Sunosuchus ’-like taxon, and will certainly bring important new information on the evolution and distribution of Goniopholididae .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.