
Diaphorocetus ortegai, Lambert & Collareta & Benites-Palomino & Merella & Muizon & Bennion & Urbina & Bianucci, 2023
|
publication ID |
https://doi.org/ 10.5252/geodiversitas2023v45a22 |
|
publication LSID |
urn:lsid:zoobank.org:pub:74B9ACDB-AEF0-40C4-97C8-D12D071A4CD4 |
|
DOI |
https://doi.org/10.5281/zenodo.10249977 |
|
persistent identifier |
https://treatment.plazi.org/id/49F43A5D-ECEB-4D12-AC7A-67FE04881EF4 |
|
taxon LSID |
lsid:zoobank.org:act:49F43A5D-ECEB-4D12-AC7A-67FE04881EF4 |
|
treatment provided by |
Plazi |
|
scientific name |
Diaphorocetus ortegai |
| status |
sp. nov. |
Diaphorocetus ortegai n. sp.
( Figs 2-6 View FIG View FIG View FIG View FIG View FIG )
urn:lsid:zoobank.org:act:49F43A5D-ECEB-4D12-AC7A-67FE04881EF4
Physeteroidea indet. – Di Celma et al. 2018: main map (supplemental material file #3).
cf. Diaphorocetus sp. – Bianucci et al. 2018b: 262.
cf. Diaphorocetus sp. – Bianucci & Collareta 2022: 36.
DIAGNOSIS. — Diaphorocetus ortegai n. sp. is a small- to medium-sized physeteroid (BZW c. 500 mm) differing from Diaphorocetus poucheti by the following cranial characters: smaller cranial dimensions; higher number of alveoli per length unit along the alveolar groove (5.2-5.4/ 100 mm), most likely resulting in a higher tooth count (>18 per upper toothrow); right premaxillary foramen located posterior to the level of the antorbital notch; and absence of a large dorsal infraorbital foramen in the posterodorsal region of the right maxilla, nearly in line with the nuchal crest.
HOLOTYPE. — MUSM 3246 , a partly complete cranium lacking the distal tip of the rostrum, lateral portions of the neurocranium (including most of the supraorbital processes and part of the zygomatic processes of the squamosals), most of the occipital shield, teeth, and ear bones ( Figs 2-6 View FIG View FIG View FIG View FIG View FIG ; Appendix 2 for 3D model).
TYPE LOCALITY. — Ullujaya , East Pisco Basin, southern coast of Peru. Geographic coordinates: 14°35’06.02”S, 75°38’06.6”W ( Fig. 1 B View FIG ). GoogleMaps
TYPE HORIZON AND AGE. — Chilcatay Formation, Ct1 allomember, Ct1a facies association. The age of the type horizon is constrained to the 18.8-18.3 Ma interval (see above for more details; Fig. 1D View FIG ).
ETYMOLOGY. — The eponymous name honours Prof. Dr. Hernán Ortega, Peruvian ichthyologist and former director of the Museo de Historia Natural, Universidad Nacional Mayor de San Marcos, for his continuous support of palaeontology in Peru, especially during the early years of the Departamento de Paleontología de Vertebrados at the Museo de Historia Natural.
DESCRIPTION
General cranial morphology
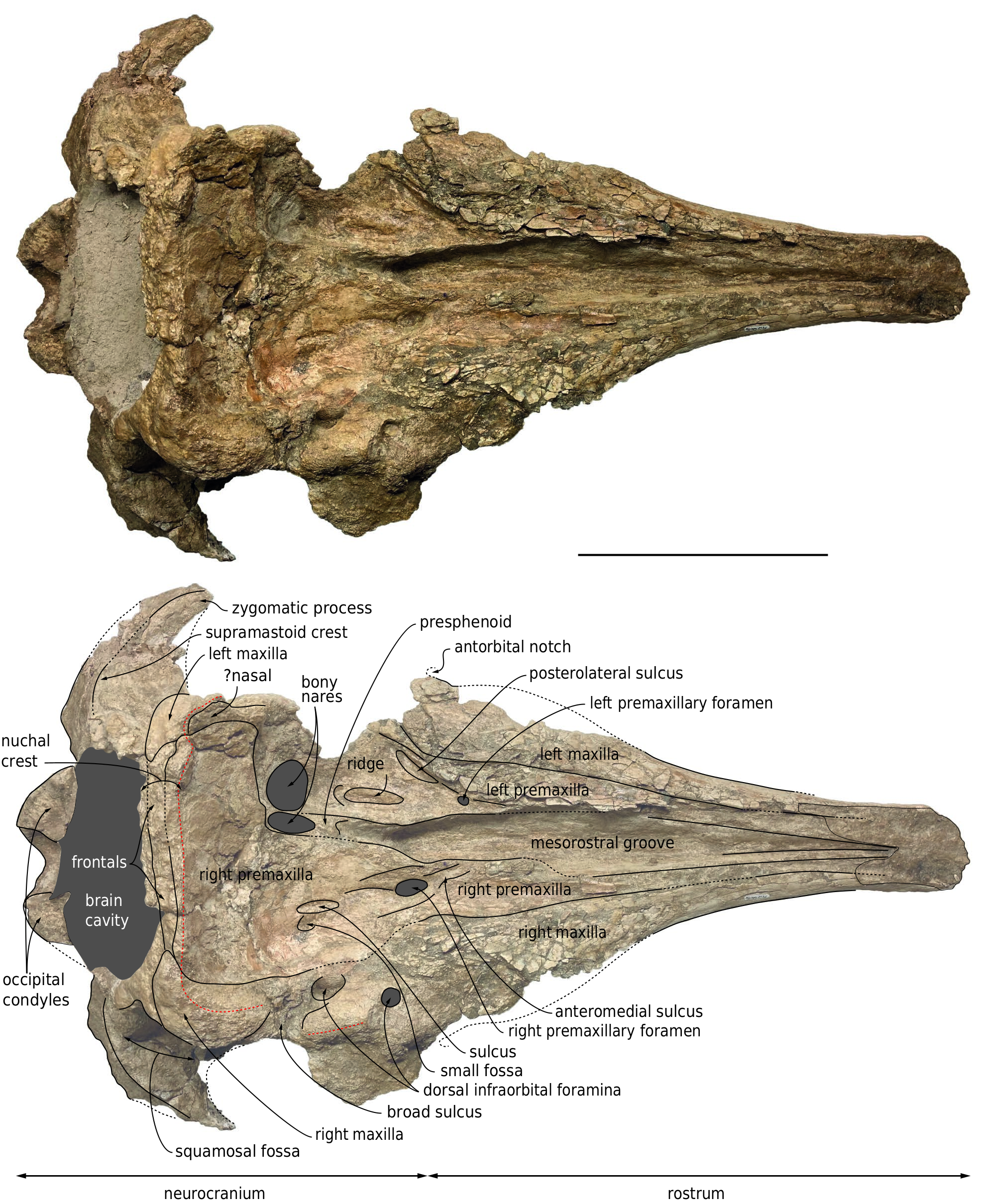
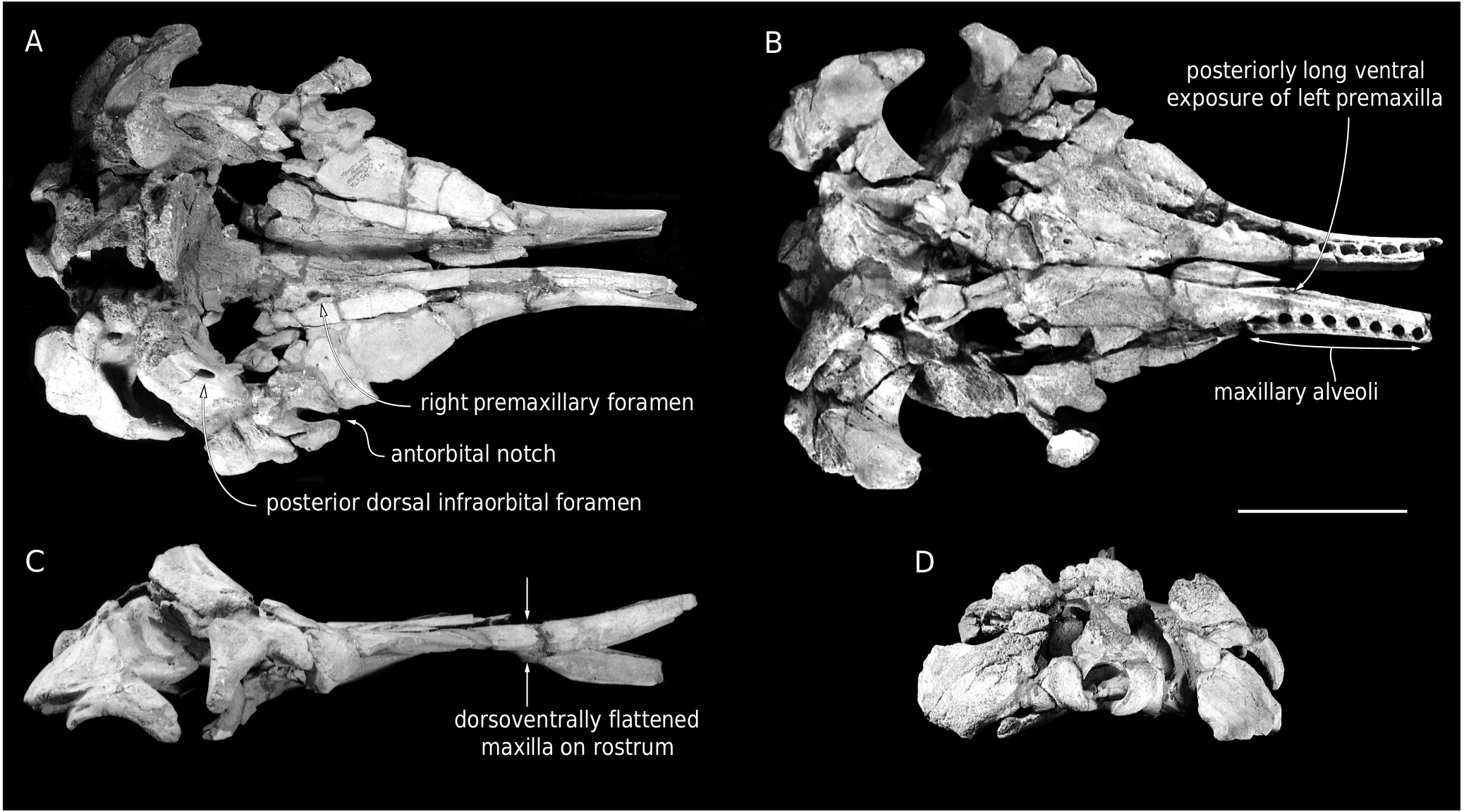
In addition to missing the anterior tip of the rostrum, lateral and posterodorsal portions of the neurocranium, MUSM 3246 is characterized by damaged surfaces, with many post-mortem fractures, especially on the dorsal surface of the rostrum and walls of the supracranial basin, as well as the zygomatic arches. The dorsal surface of the anterior part of the rostrum has been heavily abraded, due to prolonged exposure to weathering in the present-day desert environment. In addition to the truncation of the rostrum, part of the premaxillae has been lost medially along the dorsal portion. Bone surfaces are better preserved ventrally, but a thin layer of sediment has been retained in several places, e.g. in the right orbital region and posterolateral parts of the palate, due to low visual contrast with the underlying spongy bones.
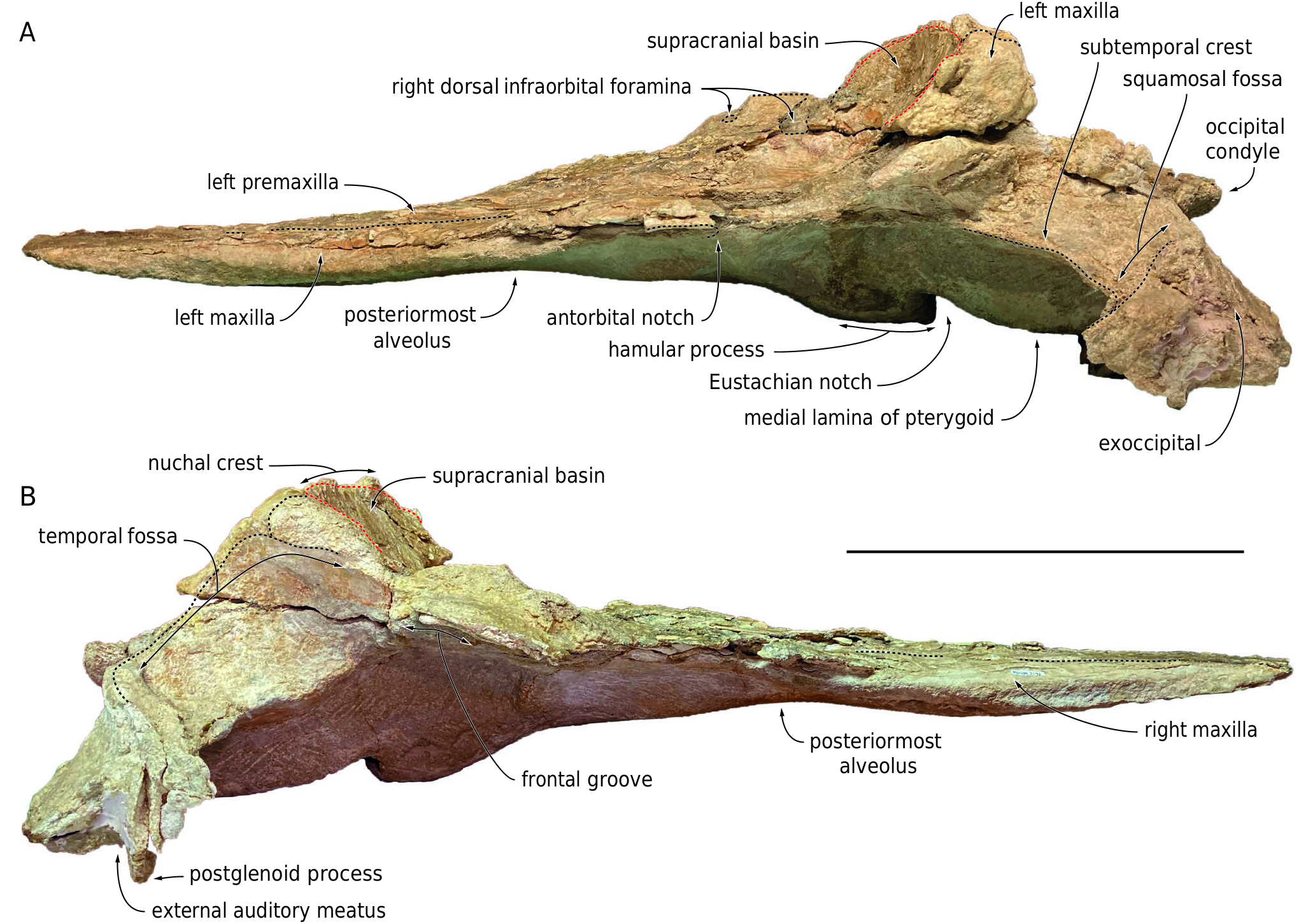
Considering the closed (but not fused) sutures for all the preserved cranial bones, their general robustness, the well-defined mandibular dental alveoli, and the presence of embrasure pits along the palatal surface of the right maxilla (see below), the ontogenetic stage of the individual represented by this cranium is interpreted as adult or subadult. With an estimated BZW of 500 mm, this medium-sized physeteroid was close in cranial width to specimens referred to Rhaphicetus valenciae (BZW = 505 mm in holotype), Placoziphius duboisi Van Beneden, 1869 (BZW roughly estimated at 490 mm in holotype), and Orycterocetus crocodilinus Cope, 1867 (BZW = 485 mm in USNM 22926). Using the BZW equation of Pyenson & Sponberg (2011) for stem physeteroids, body length of MUSM 3246 is estimated to 4.54 m. Most cranial dimensions are smaller than in the holotype of Diaphorocetus poucheti ( Table 1 View TABLE ; Paolucci et al. 2020) and, to an even greater extent, Cozzuoliphyseter rionegrensis ( Paolucci et al. 2021) . Since the rostrum is truncated at a level where its lateral margins are parallel and distant from each other ( Figs 2 View FIG ; 3 View FIG ), it was markedly longer than the neurocranium. The proximal portion of the rostrum is much wider than high in cross section (defined as the platyrostrine condition). In lateral view, the ventral outline of the rostrum is moderately convex in its anterior preserved portion, which is slightly anterodorsally recurved ( Fig. 4 View FIG ); it becomes distinctly concave in its most proximal region, thus drawing a typical sigmoid shape.
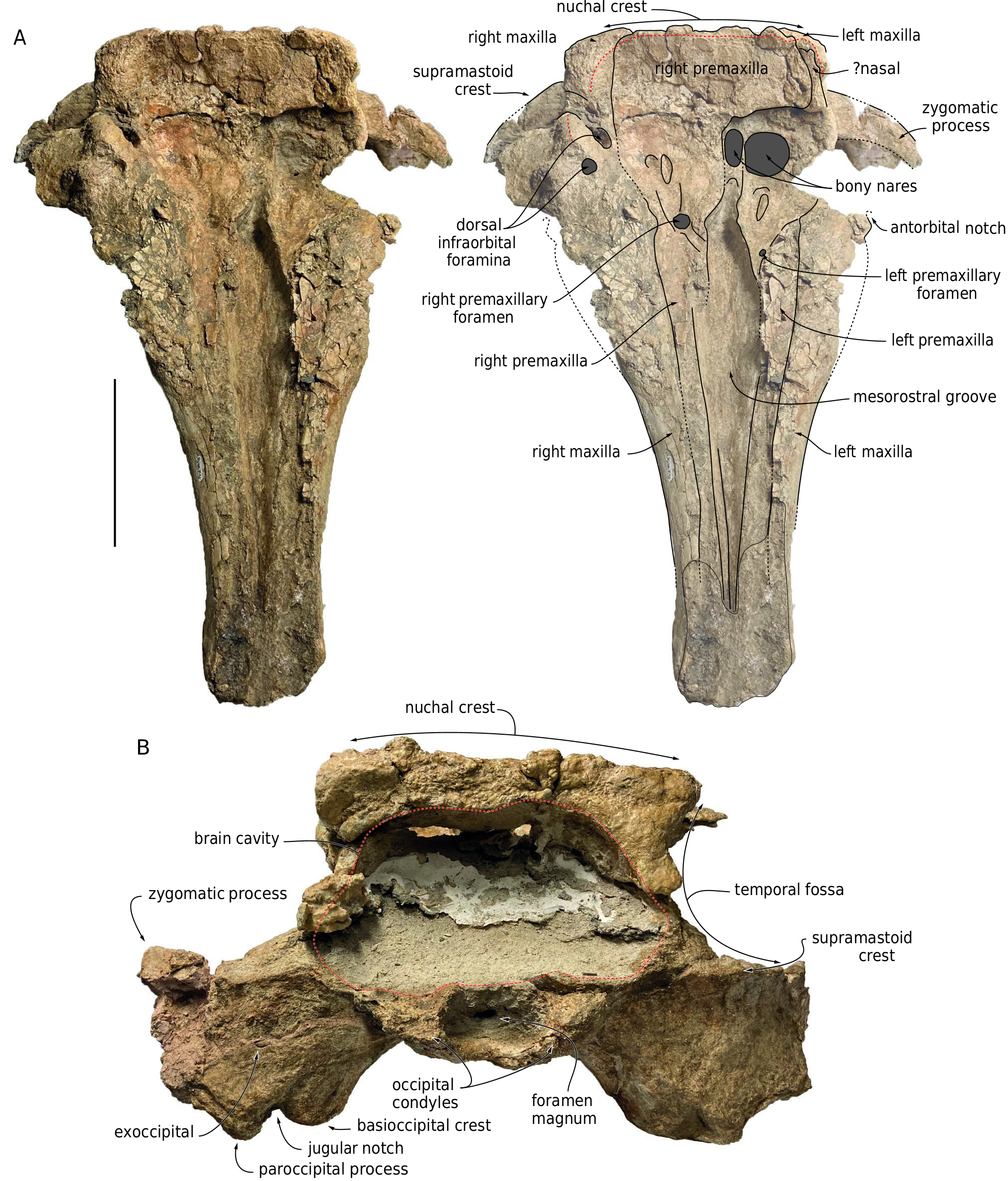
With the nuchal crest being rectilinear in dorsal view, the supracranial basin has a sub-rectangular posterior outline, as seen in Aulophyseter morricei Kellogg, 1927 , C. rionegrensis , D. poucheti , and, to a lesser extent, R. valenciae (where it is more rounded on the left side). The basin ends posteriorly before the level of the squamosal fossa, and laterally before the level of the antorbital notch (outline of posteromedial margin of left antorbital notch partly preserved in lateral view) ( Figs 2 View FIG ; 5 View FIG ; 6 View FIG ). The posterior wall of the basin is low, with the nuchal crest reaching barely higher dorsally than the temporal fossa, 75 mm above the dorsal margin of the right bony naris. This corresponds to a basin that is much less dorsally extended than in e.g. Acrophyseter spp. , Orycterocetus crocodilinus , and Physeter macrocephalus Linnaeus, 1758 . The dorsomedial elevation of the medial margin of the left premaxilla just anterior to the left premaxillary foramen suggests that the supracranial basin did not extend for a long distance along the rostrum base, a condition shared with most other extinct physeteroids. This interpretation is further supported by the proportions of the rostrum, which is tapered in its proximal region, and as such, much narrower than in Livyatan melvillei (Lambert, Bianucci, Post, Muizon, Salas-Gismondi, Urbina & Reumer, 2010) and P. macrocephalus for its preserved anterior part. As in all other physeteroids, the left bony naris is much larger than the right (ratio between transverse width of right and left naris is estimated at 0.46; Table 1 View TABLE ). The right naris is displaced to the left side of the sagittal plane of the skull, in line with the posterior tract of the mesorostral groove.
The preserved anterodorsal roof of the right temporal fossa and parts of the supramastoid crests indicate that the temporal fossae were voluminous, i.e., transversely broad (more than 120 mm) as well as considerably higher dorsoventrally and longer anteroposteriorly than observed in P. macrocephalus . Although most of the occipital shield is missing, the anteroposterior position of the non-deformed nuchal crest (markedly anterior to the occipital condyles) indicates that the shield was originally inclined in an anterodorsal direction, drawing an angle of about 45° with the horizontal ( Fig. 4 View FIG ).
Premaxilla
Although the dorsal surface of each premaxilla is nearly completely lost along the anterior half of the rostrum, marks of the sutures with the maxillae indicate that the premaxillae did not make the whole dorsal surface of the rostrum in that region, considering the dorsal exposure of the maxillae for a length of at least 310 mm ( Fig. 2 View FIG ). The medial edge of the left premaxilla is somewhat better preserved in the proximal portion of the rostrum, attesting to a broad dorsal opening of the mesorostral groove at the rostrum base, with a maximum opening being reached 100 mm anterior to the level of the antorbital notch. It remains unknown whether the right and left premaxillae contacted each other dorsomedially in the anterior part of the rostrum.
The left premaxillary foramen is located just anterior to the level of the antorbital notch, 8.5mm lateral to the medial margin of the premaxilla, with a transverse diameter of 7 mm ( Fig. 2 View FIG ). The foramen marks the posterolateral corner of a short (25 mm long), depressed, and transversely concave surface that may correspond to the prenarial triangle of many non-physeteroid odontocetes. Anterior to this region, the heavily fragmented dorsal surface of the premaxilla is slightly concave transversely for at least 100 mm. Posterior to the premaxillary foramen, the left premaxilla widens markedly. Its dorsal surface is transversely concave, forming a broad premaxillary sac fossa that is divided by a thick longitudinal ridge for about 43 mm. Medial to this ridge, the fossa is even more concave, with a medial edge raising dorsomedially. At this level, the left premaxilla remains distant from the right, leaving the posterior part of the mesorostral groove dorsally open.Twenty-nine millimeters posterolateral to the left premaxillary foramen, a moderately deep sulcus extends obliquely for at least 44 mm and ends parallel to the lateral margin of the premaxilla, being reminiscent of the posterolateral sulcus of non-physeteroid odontocetes (see Lambert et al. 2020 for a similar interpretation in Rhaphicetus valenciae ). The anterior margin of the left bony naris is located shortly posterior to the end of the longitudinal ridge. Posterolaterally, the left premaxilla is incomplete, thus preventing from estimating its extent along the lateral wall of the supracranial basin.
The dorsal surface of the right premaxilla is transversely concave for at least 160 mm anterior to the right premaxillary foramen ( Figs 5 View FIG ; 6 View FIG ), a marked difference with the convex premaxilla of R. valenciae in this region. The right premaxillary foramen is located 35 mm more posteriorly than the left, shortly behind the proposed level of the antorbital notch; in Diaphorocetus poucheti , this foramen is located more anteriorly. The right premaxillary foramen is more than twice larger (transverse diameter 16 mm) than the left.It is followed anteriorly for 36 mm by a well-defined, broad anteromedial sulcus, a feature that was not observed in R. valenciae . Another sulcus extends posteriorly from the premaxillary foramen; it is shorter, narrower, and shallower than in the holotype of R. valenciae . Smaller depressions mark the right premaxillary sac fossa more posteriorly, including a short and narrow sulcus (29 mm long and 8 mm wide) located medial to a small fossa. Posteromedial to the premaxillary foramen, the surface of the right premaxilla is transversely convex, with a medial edge turning posteromedially towards the presphenoid.
The suture between the right maxilla and premaxilla is difficult to follow in the supracranial basin due to damage in that region; it is estimated that the lateral margin of the premaxilla roughly reaches the medial limit of the largest dorsal infraorbital foramen (see below), at about half the height of the lateral wall of the supracranial basin, having at that level a maximum width of 97 mm. From there, the premaxilla raises onto the posterior wall of the basin, where it contributes to the anteroposteriorly thick nuchal crest, and where the dorsal-most portion of its anteriorly facing surface is vertical. Covering most of the posterior wall of the supracranial basin, the right premaxilla extends far towards the left part of the skull, reaching beyond the lateral margin of the left bony naris ( Figs 2 View FIG ; 5 View FIG ), as seen for example in Cozzuoliphyseter rionegrensis ( Paolucci et al. 2021) .
On the ventral surface of the rostrum, the two premaxillae are exposed in the anterior part of the median trough, displaying an asymmetric pattern ( Fig. 3 View FIG ). The left premaxilla appears 45 mm before the anterior end of the vomer, whereas the right premaxilla remains hidden until a level 50 mm anterior to the vomer, from where it broadens to eventually reach a width similar to that of the left premaxilla at the preserved tip of the rostrum. A similar asymmetry is observed in D. poucheti , P. duboisi , and C. rionegrensis .
Maxilla
The anterior tip of the maxillae is missing, and the abraded dorsal surface makes it difficult to estimate the length of the missing part. Each maxilla is exposed dorsally along the whole preserved length of the rostrum; this exposure widens only slightly for the first 250 mm, followed by a more pronounced widening linked to the broadening of the rostrum towards its base ( Figs 2 View FIG ; 5 View FIG ). Using the partly preserved posteromedial wall of the left antorbital notch as a landmark, it is probable that a moderately developed maxillary flange was originally present. In lateral view, the maxillary part of the rostrum is surprisingly thin dorsoventrally for the preserved anterior two thirds of its length ( Fig. 4 View FIG ), contributing to the platyrostrine (dorsoventrally flattened) aspect of the rostrum: for example, the maxilla is 32 mm thick 250 mm anterior to the antorbital notch in MUSM 3246, whereas it is 63 mm thick at the same level in the holotype of Rhaphicetus valenciae . A similar dorsoventrally flattened maxillary portion of the rostrum is seen in Diaphorocetus poucheti ( Fig. 7 View FIG ; Paolucci et al. 2020) and in the less closely related Zygophyseter varolai ( Bianucci & Landini 2006) .
On the left side, anteromedial to the antorbital notch, the dorsal surface of the maxilla is transversely convex, whereas on the right side it is concave ( Figs 5 View FIG ; 6 View FIG ), as in R. valenciae . Only the right maxilla is partly preserved in the supraorbital region. It displays two dorsal infraorbital foramina ( Figs 2 View FIG ; 5 View FIG ). With a transverse diameter of 9.5 mm and a circular outline, the first such foramen is just posterior to the level of the antorbital notches, being placed at the anterior end of the thick crest that makes the lateral margin of the supracranial basin over the right orbit. The second foramen is larger (anteroposterior diameter estimated at 24 mm), oval in outline, and located 32 mm more posteromedially, along the medial surface of the thick crest. It is followed posterolaterally by a broad sulcus. It cannot be excluded that smaller foramina went unnoticed due to preservation issues on the right side. A large dorsal infraorbital foramen is located more posterodorsally on the right maxilla of the holotype of D. poucheti , nearly in line with the nuchal crest. No dorsal infraorbital foramen is preserved on the left side ( Fig. 7 View FIG ).
The posterolateral corners of the supracranial basin are mostly made of the maxillae and markedly thickened. Both maxillae extend posteromedially along the nuchal crest for a short distance. Each maxilla ends before the mediolateral level of the corresponding premaxillary foramen, the two maxillae being approximately 120 mm apart.
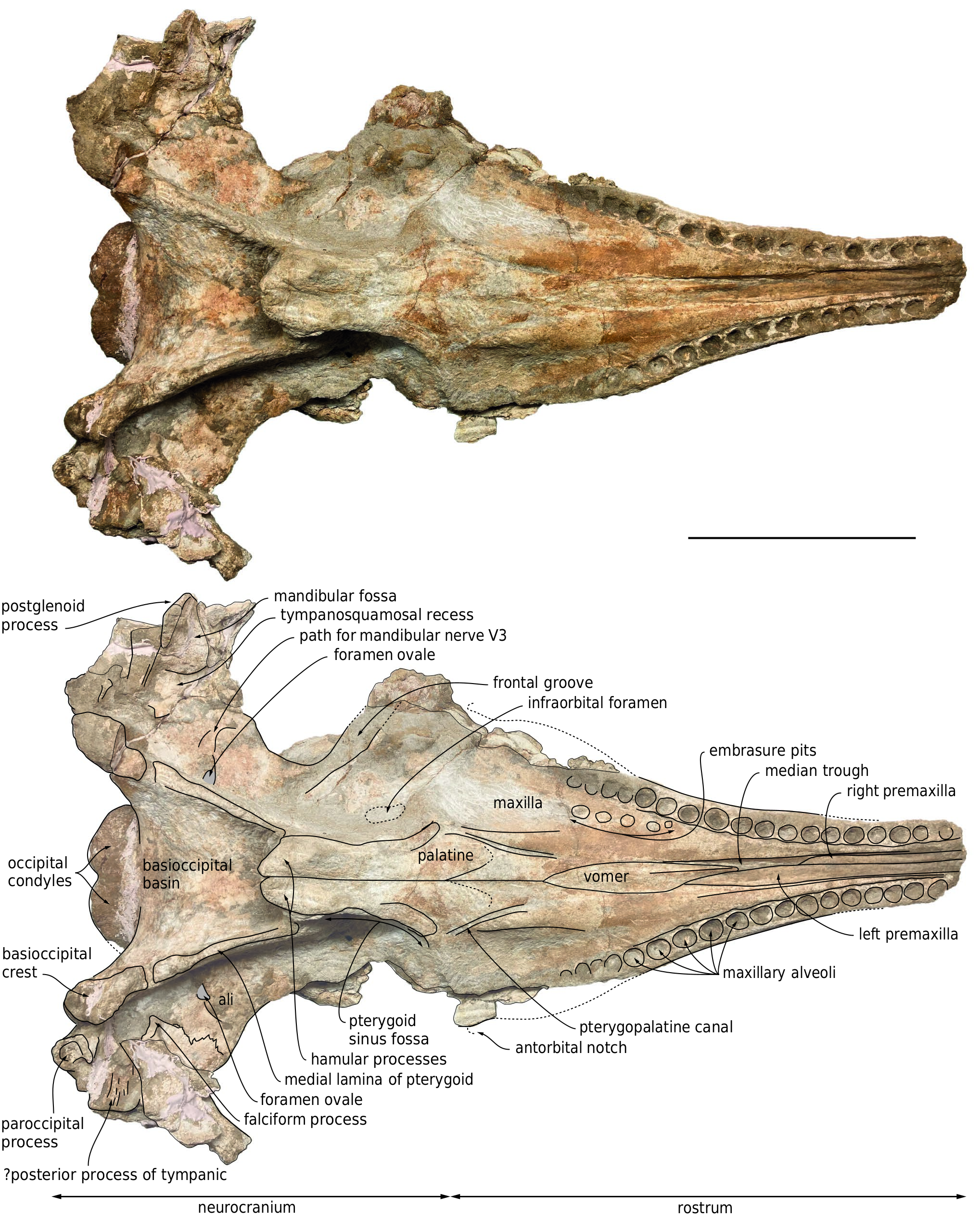
In ventral view, the left and right maxillae display 18 dental alveoli over a length of 336 mm and 347 mm, respectively ( Fig. 3 View FIG ). In addition to maxillary and presumably premaxillary alveoli from the missing anterior section of the rostrum, a few more, shallow alveoli may have been present posteriorly, along the damaged lateral margins of the basal portion of the rostrum. The number of alveoli per 100 mm of alveolar groove length is greater than in D. poucheti . The alveolar groove of Placoziphius duboisi probably ended more anteriorly compared to MUSM 3246, with shallower posterior alveoli. The right and left anteriormost alveoli are 36.5 mm apart from each other, whereas the right and left posteriormost alveoli are 166 mm from each other, and the last preserved posterior alveolus is 76 mm anterior to the antorbital notch on the left side. The preserved alveoli are well-defined, roughly circular, and deep. They range in transverse diameter from 13 to 18 mm, with the largest occurring at about two thirds of the length of the alveolar groove (13rd to 14th alveoli, numbered from the first preserved anterior alveolus), and the smallest being in the first half (from 4 th to 9 th along the left alveolar groove). Interalveolar septa range in length from 3 to 18 mm, but most of the septa fall in an interval between 3 and 5.5 mm, with no general trend along the alveolar groove.
Medial to the 17th to 12th right alveoli, five moderately well-defined, subcircular depressions most likely correspond to embrasure pits. The posteriormost such pit is the deepest, with a central part that is 22.5 mm medial to the alveolar groove, and subsequent more anterior pits get gradually shallower. Similar depressions interpreted as embrasure pits have been described medial to the posterior maxillary alveoli in other physeteroids (e.g., Acrophyseter deinodon , D. poucheti , and Orycterocetus crocodilinus ), several other odontocetes, and basilosaurids ( Lambert et al. 2017 and references therein; Paolucci et al. 2020). Either the embrasure pits were shallower, or completely absent on the left side, as seen in several pomatodelphinine platanistids ( Kellogg 1959; Lambert 2006). The palate is robust, featuring a dorsoventrally thick, 85-mm-wide ventromedial portion of the maxillae with a roughly flat ventral surface, being separated from the transversely concave lateral portions by two parallel crests. Medial to each crest, a deep oblique sulcus is directed anteromedially.The posterior part of each sulcus corresponds to the pterygopalatine canal, which is probably exposed ventrally due to poor preservation of the ventral surface of the maxilla and palatine in this area, whereas the anterior part most likely exited from an unpreserved major palatine foramen.
right premaxillary foramen maxillary alveoli antorbital notch dorsal infraorbital foramen
D
Vomer
Ventrally, the vomer is exposed along the median trough for 180 mm, starting from a level 56 mm anterior to the antorbital notches, and reaching a maximum width of 28 mm ( Fig. 3 View FIG ). From this level forward, the ventral surface of the vomer becomes keeled. Posterior to the choanae, the sutures of the vomer with other bones in the basioccipital basin could not be clearly detected due to damaged bone surfaces and partial covering by sediment.
Palatine and pterygoid
Part of the maxilla-palatine suture is visible on the right side, leaving the sagittal plane of the skull in an anterolateral direction from a level 35 mm posterior to the antorbital notch ( Fig. 3 View FIG ). The anterior tip of the right palatine is about 25 mm anterior to the level of the antorbital notch. The anterolateral portion of each pterygoid can be outlined as a narrow (15 mm wide) plate participating to the anteromedial edge of the pterygoid sinus fossa. This edge rapidly becomes poorly defined lateral to the anterior tip of the pterygoid, which is located posterior to the level of the antorbital notch, ending 55 mm posterior to the apex of the palatine. Ventromedially defining the broad pterygoid sinus fossae, the hamular processes of the pterygoids are massive, with a maximum joined width of 58 mm and a depressed median region. The posterior tips of the right and left pterygoids are joined and project for 25 mm posterior to the bottom of the Eustachian notches. The medial lamina of the pterygoid makes a long portion (117 mm on the left side) of the lateral wall of the basioccipital basin. Being slender anteriorly, it gradually thickens posterolaterally to reach a transverse thickness of 21 mm.
Presphenoid
In dorsal view, the presphenoid is only visible for a short distance between the posterior end of the mesorostral groove and the right bony naris ( Figs 2 View FIG ; 5 View FIG ). Details of the relationships with the vomer and premaxillae cannot be distinguished.
Nasal
Along the left lateral margin of the right premaxilla, on the left posterolateral corner of the supracranial basin, a narrow and thin plate of bone is tentatively interpreted as part of a single nasal bone ( Figs 2 View FIG ; 5 View FIG ).
Frontal
Dorsally, apart from the exposure of part of the right supraorbital process due to removal of the overlying maxilla, frontals are thought to appear only along the thick nuchal crest, in the section separating the two maxillae, between the left premaxilla and small fragments of the supraoccipital ( Fig. 2 View FIG ). The only feature of the frontal that is visible in ventral view is the narrow medial portion of the right frontal groove, which is obliquely oriented to form an angle of about 45° with the sagittal plane of the cranium ( Fig. 3 View FIG ).
Occipital shield and brain cavity
No part of the supraoccipital is preserved, leaving the brain cavity open ( Figs 2 View FIG ; 5 View FIG ). The latter has a preserved maximum width of 210 mm, in its lower half; it is markedly wider than high, a condition that most likely reflects the shape of the brain, as seen for example in Kogia sima Owen, 1866 and Physeter macrocephalus ( Flower 1867; Ries & Langworthy 1937; Marino et al. 2003). The minimum distance between the temporal fossae across the occipital region makes less than 43 % of the bizygomatic width ( Table 1 View TABLE ), corresponding to a relatively narrow occipital shield, laterally margined by transversely broad temporal fossae. The shield is dorsoventrally low, as testified by the low position of the nuchal crest (see above), hence a stocky posterior outline of the skull.
Squamosal
In dorsal view, the preserved part of each zygomatic process is directed anterolaterally, before gradually turning to a more anterior direction ( Fig. 2 View FIG ). Considering the direction of the anterior part of the zygomatic process in other physeteroids with a long temporal fossa, it is estimated that a large part of each zygomatic process (at least 90 mm on the right side) is missing. The supramastoid crest raises posterodorsomedially,as in other physeteroids ( Fig.4 View FIG ).The right postglenoid process is partly preserved,making a slightly obliquely directed thin plan (thickness 8-9 mm) and reaching farther ventrally than the posttympanic process, as in Diaphorocetus poucheti . In ventral view,the left falciform process has a triangular outline, with a dorsoventrally thin ventromedial tip ( Fig.3 View FIG ).Part of the tympanosquamosal recess is better seen on the right side, medial to the postglenoid process, and extending anterolaterally along the damaged mandibular fossa.
Alisphenoid
Finely preserved on the left side, the ventral suture between alisphenoid and squamosal zigzags in an anterolateral direction, reaching the subtemporal crest some distance anteromedial to the anterior most edge of the squamosal fossa ( Fig. 3 View FIG ). Filled with hardened sediment on both sides, the foramen ovale is followed laterally by a wide and shallow path for the mandibular nerve (V3), which gradually turns more anteriorly, disappearing before the subtemporal crest.Anterior to this path, the ventral surface of the alisphenoid is approximatively flat, sloping anterodorsolaterally.
Exoccipital
The posterior surface of the exoccipital slopes posteroventrally, with an angle of 45-50° with the horizontal plane ( Fig. 4 View FIG ). Only the ventral part of the occipital condyles is preserved, being lower than the posteroventral margin of the temporal fossae ( Fig. 5 View FIG ). The condyles protrude moderately from the neurocranium, reaching posteriorly just beyond the ventral margin of the exoccipitals. The ventral surface of the paroccipital process is deeply excavated, with a laterally directed broad groove (better preserved on the left exoccipital) ( Fig. 3 View FIG ). Anterolateral to the left paroccipital process, a broad and dorsoventrally thin piece of bone ventrally marked by laterally directed grooves and ridges is tentatively interpreted as part of the posterior process of the left tympanic bulla.
Basioccipital
Though the basioccipital basin is anteroposteriorly long (147 mm from the posterior tip of the hamular processes), the contribution of the basioccipital crests to its dorsolateral walls is relatively limited (see medial lamina of pterygoid above) ( Fig. 3 View FIG ). The dorsal surface of the basioccipital basin is markedly anteroposteriorly convex. The posterolateral portion of each basioccipital crest is robust (maximum transverse thickness equaling 38 mm on the left side), medially defining a moderately deep and broad jugular notch.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Diaphorocetus ortegai
| Lambert, Olivier, Collareta, Alberto, Benites-Palomino, Aldo, Merella, Marco, Muizon, Christian de, Bennion, Rebecca, Urbina, Mario & Bianucci, Giovanni 2023 |
Diaphorocetus sp.
| BIANUCCI G. & COLLARETA A. 2022: 36 |
Diaphorocetus sp.
| BIANUCCI G. & COLLARETA A. & BOSIO G. & LANDINI W. & GARIBOLDI K. & GIONCADA A. & LAMBERT O. & MALINVERNO E. & MUIZON C. DE & VARAS-MALCA R. & VILLA I. M. & COLETTI G. & URBINA M. & DI CELMA C. 2018: 262 |