
Cistenides hyperborea Malmgren, 1865
|
publication ID |
https://doi.org/10.5852/ejt.2020.666 |
|
publication LSID |
lsid:zoobank.org:pub:2E0FAA1D-DA9A-4486-805F-9DA3DF928539 |
|
DOI |
https://doi.org/10.5281/zenodo.5919026 |
|
persistent identifier |
https://treatment.plazi.org/id/03B4E316-5028-FFB0-FDCD-FA82313AE269 |
|
treatment provided by |
Valdenar |
|
scientific name |
Cistenides hyperborea Malmgren, 1865 |
| status |
|
Cistenides hyperborea Malmgren, 1865 View in CoL
Figs 1D View Fig , 2D View Fig , 3D View Fig , 4 View Fig E–H, 10 View Fig – 15 View Fig
Cistenides hyperborea Malmgren, 1865: 360 View in CoL .
Pectinaria hyperborea View in CoL – Hessle 1917: 76.
Pectinaria ( Cistenides) hyperborea View in CoL – Nilsson 1928: 31. — Pettibone 1954: 312, fig. 35c-h. — Holthe 1986: 25. — Hartmann-Schröder 1996: 482.
Material examined ( 125 specimens, 11.7% of total specimens identified, in ten samples)
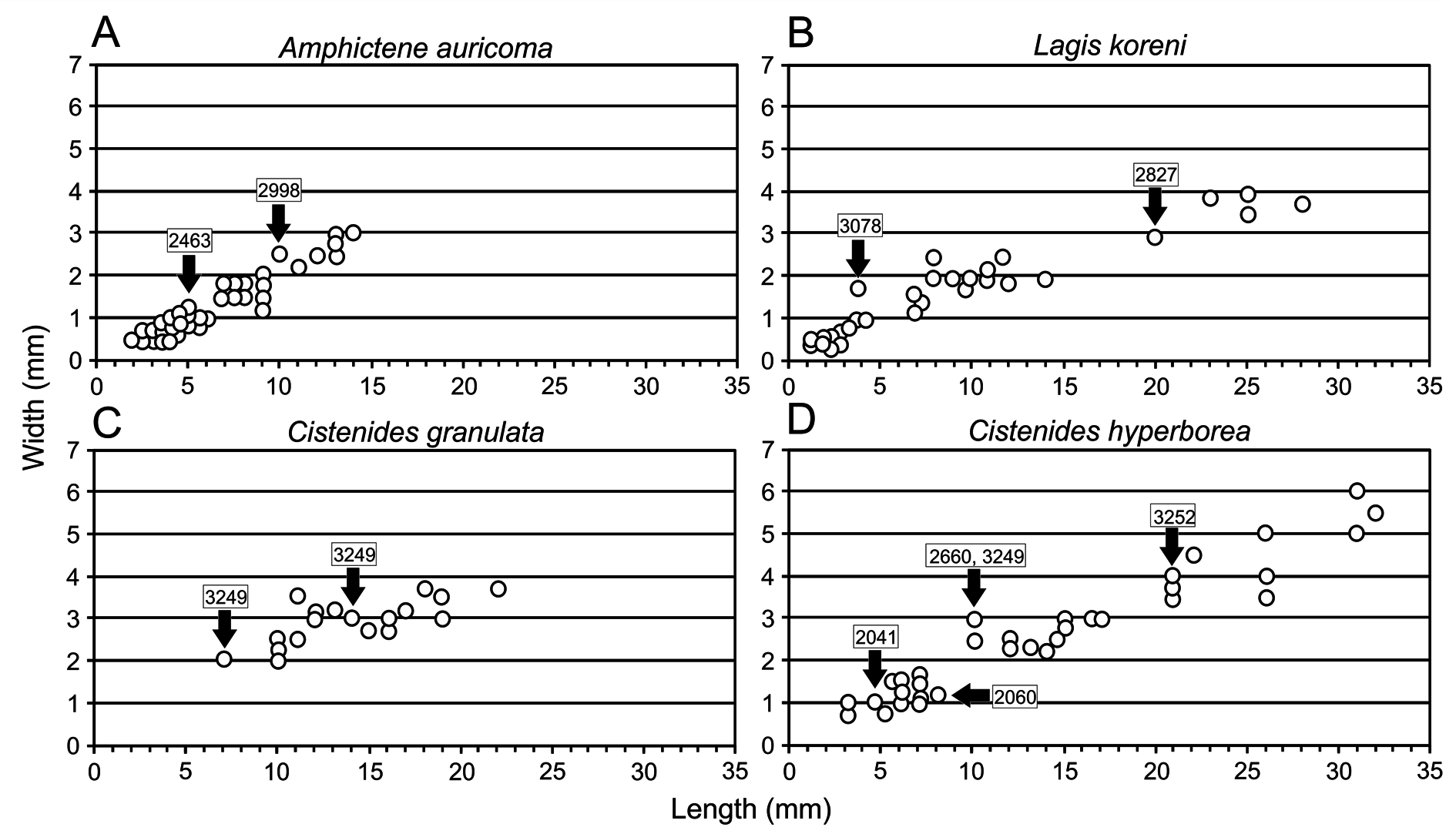
ICELAND – North coast • 1 spec.; BIOICE station 1, sample 2001; 65º21′19″ N, 13º47′61″ W; 19 Jul. 1991; 6.0ºC; salinity unknown; 26 m depth; organic silt; IINH-40468 GoogleMaps • 3 specs; BIOICE station 3, sample 2005; 65º20′88″ N, 13º18′64″ W; 19 Jul. 1991; 2.8ºC; salinity unknown; 171 m depth; sandy silt and stones; MNCN 16.01/18015 GoogleMaps • 1 spec.; BIOICE station 29, sample 2041; 65º48′61″ N, 14º34′01″ W; 24 Jul. 1991; 2.8ºC; salinity unknown; 129 m depth; sediment unknown; IINH-40470 GoogleMaps • 54 specs; BIOICE station 2, sample 2060; 66º00′61″ N, 17º31′78″ W; 2 Jul. 1992; temperature unknown; salinity unknown; 48 m depth; soft sediment; IINH-40471 • 3 specs; BIOICE station 3, sample 2064; 66º02′33″ N, 17º32′21″ W; 2 Jul. 1992; 5.4ºC; 34.69 ppm; 102 m depth; mud; IINH-40472 GoogleMaps • 16 specs; BIOICE station 16, sample 2097; 66º36′92″ N, 18º14′42″ W; 5 Jul. 1992; 4.88ºC; 34.89 ppm; 110 m depth; sand and stones; IINH-40473 • 17 specs; BIOICE station 17, sample 2660; 67º14′41″ N, 15º28′42″ W; 15 Jul. 1994; 2.69ºC; 34.90 ppm; 277 m depth; sandy silt; IINH-40474 GoogleMaps • 5 specs; BIOICE station 5, sample 3108; 67º45′48″ N, 18º30′85″ W; 21 Aug. 1999; -0.21ºC; 34.88 ppm; 328 m depth; sediment unknown; IINH-40475 GoogleMaps • 23 specs; BIOICE station 23, sample 3249; 65º50′34″ N, 12º01′27″ W; 14 Jul. 2001; 1.92ºC; 34.87 ppm; 192 m depth; sediment unknown; IINH-40476 GoogleMaps • 24 specs; BIOICE station 24, sample 3252; 65º45′94″ N, 12º16′39″ W; 14 Jul. 2001; 1.48ºC; 34.86 ppm; 232 m depth; sediment unknown; IINH-40477 GoogleMaps .
Occurrence
From off northern and north-eastern coast of Iceland ( Fig. 1D View Fig ). Depth range: 26 to 328 m; bottom temperature range: -0.21 to 6.00°C ( Fig. 2D View Fig ). Water mass/es: NSAIW, CW and MNAW.
Remarks
Cistenides hyperborea – type locality: Greenland and Spitzbergen ( Holthe 1986) – is a poorly known species; the original description does not indicate the number of rows of teeth present on the uncini, although 1 to 3 rows have previously been mentioned for specimens assigned to this taxon ( Hutchings & Peart 2002). Because type material was never designated ( Holthe 1986), Hutchings & Peart (2002) suggested that specimens from the type locality need to be re-examined to ascertain the generic identity of this species.
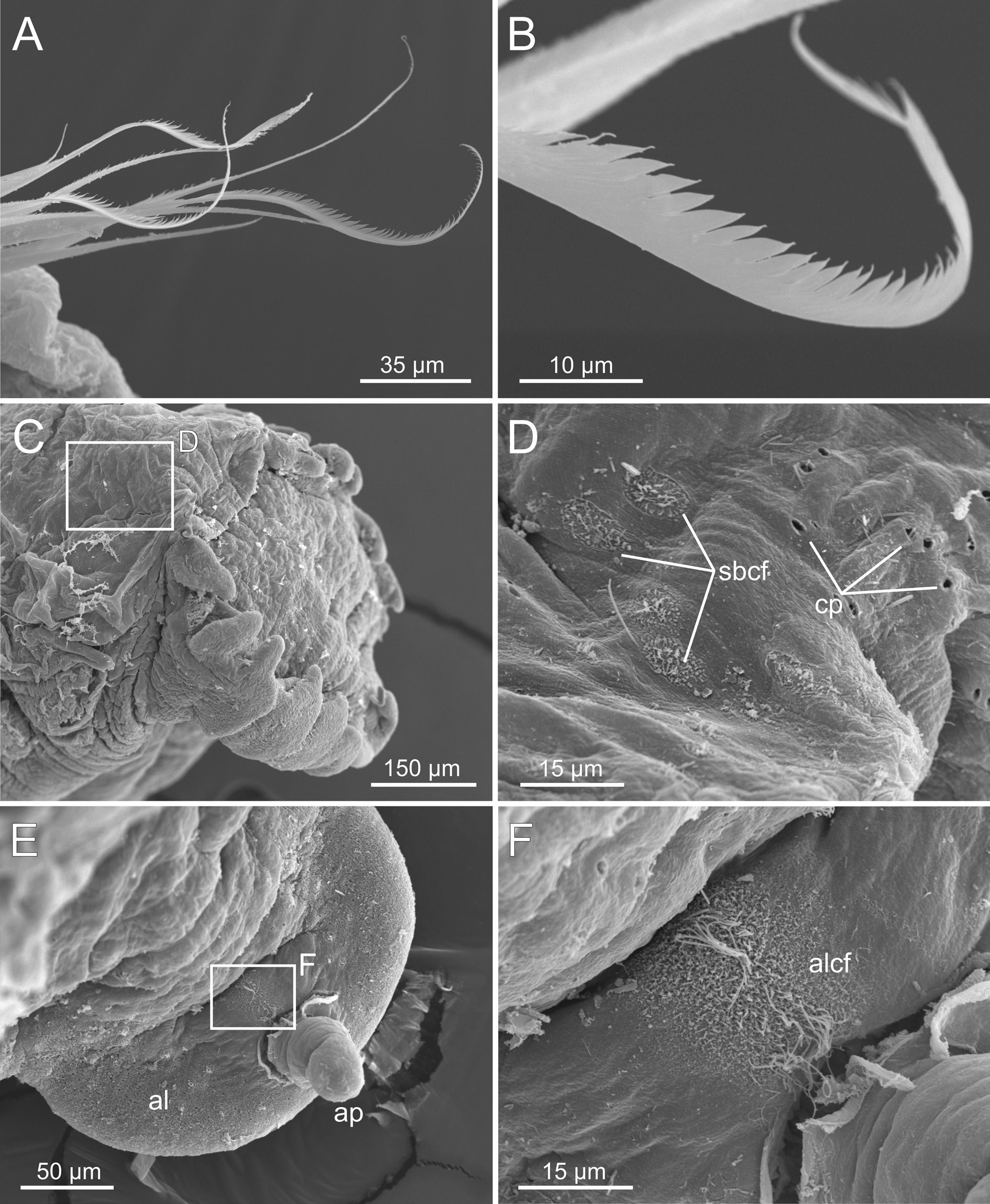
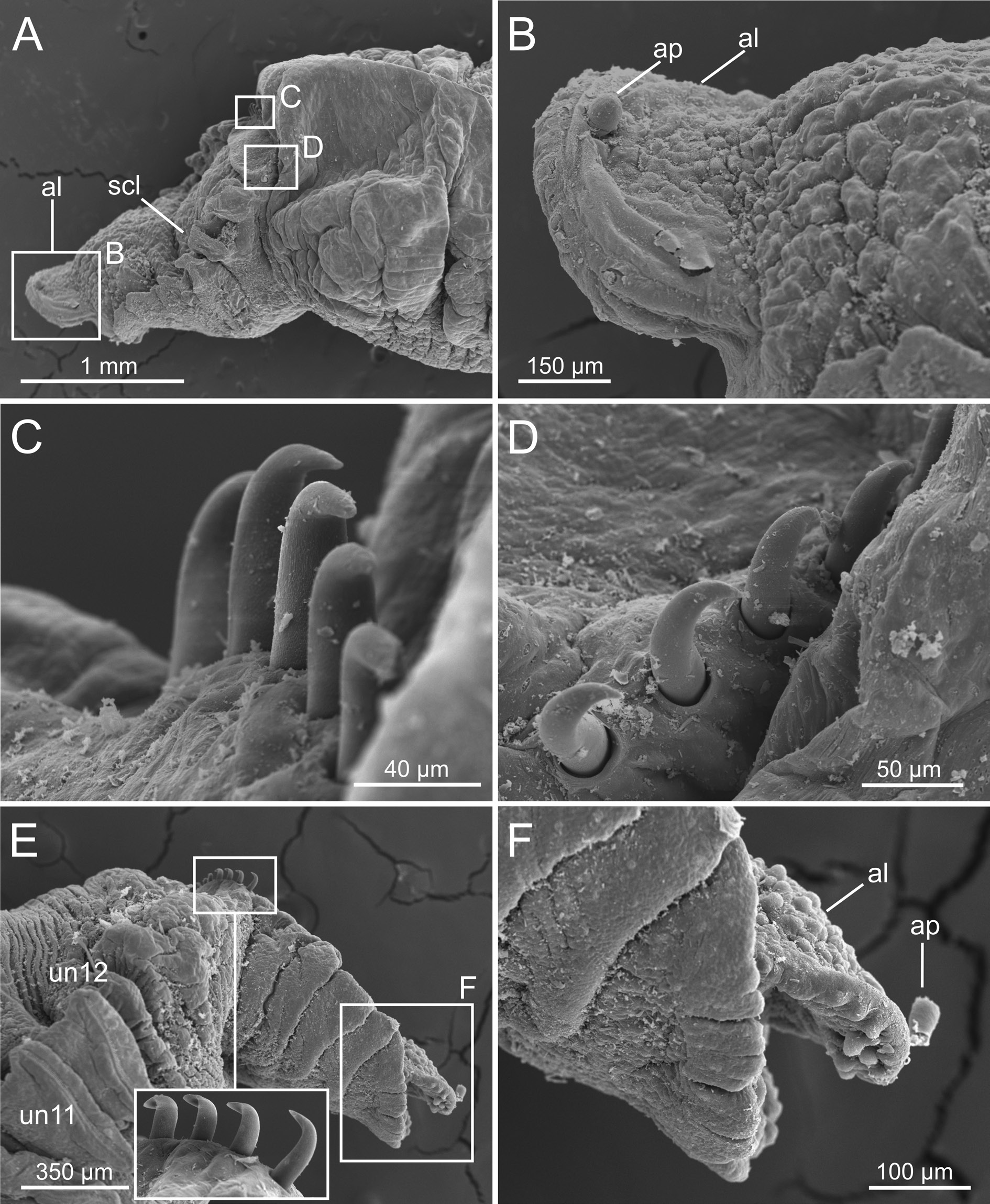
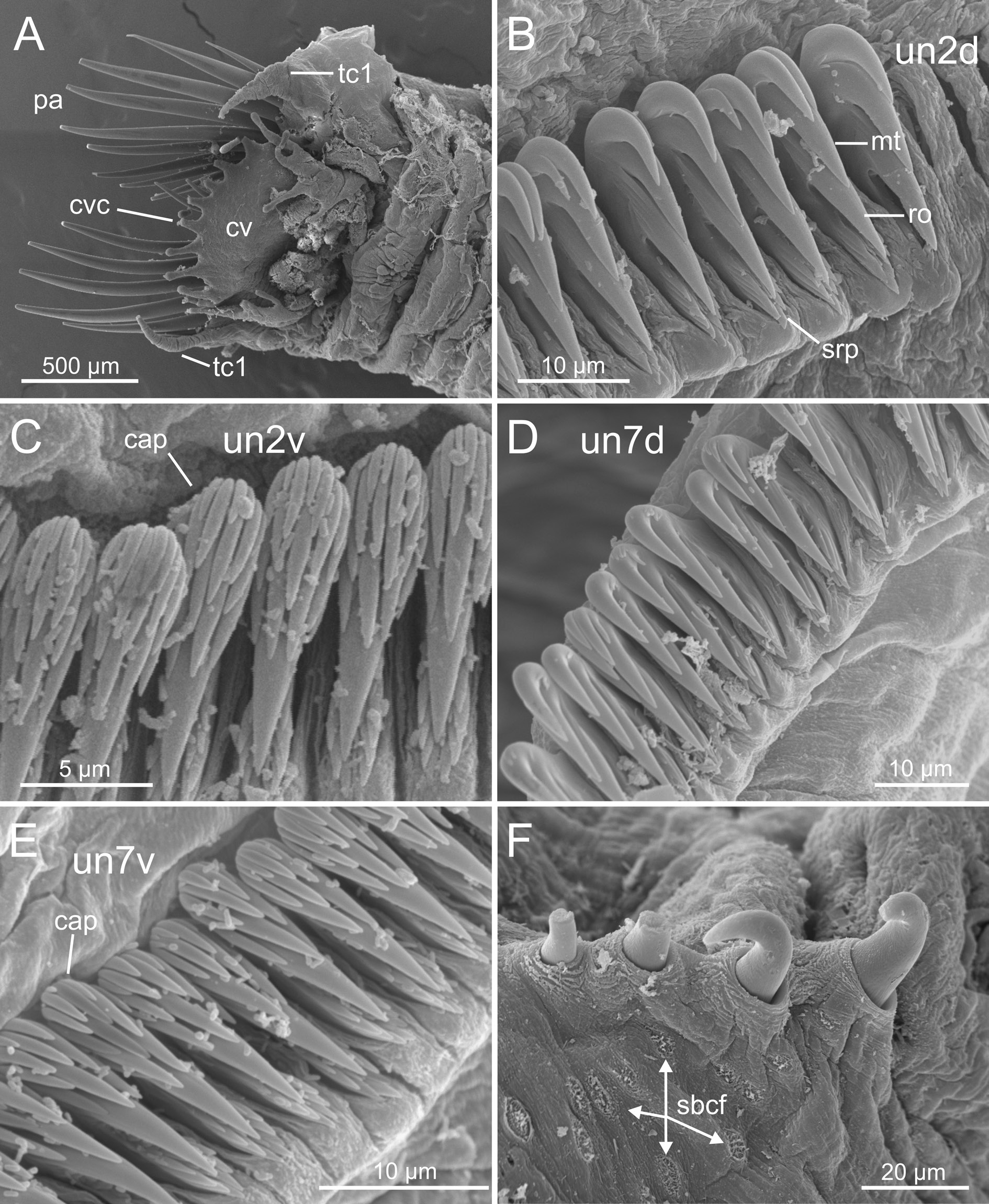
The examination of the Icelandic specimens here identified as C. hyperborea , covering a wide range of body sizes, shows a considerable variation in the shape of the neuropodial uncini, contrary to the expected, according to the description of the species. BIOICE specimens agree with previous descriptions (e.g., Pettibone 1954; Holthe 1986; Hutchings & Peart 2002) in: 1) having a smooth dorsal brim and cephalic veil free from operculum, provided with several well-defined cirri ( Fig. 10A View Fig ); 2) the relation between chaetigers with notochaetae versus those with also ventral uncini is 17/12; 3) the paleae are long, numerous, straight and with pointed tips ( Figs 10A View Fig , 12A View Fig ); 4) the scaphe has small lateral lobes ( Fig. 11A View Fig ) with a short and rounded anal lobe and anal papilla ( Fig. 11B View Fig ), and scaphal hooks are distally bent ( Fig. 11 View Fig C–E). On the other hand, the aforementioned characters show, however, differences related to the size of the specimen. For instance, medium-sized and small individuals show a strait scaphe border with a verrucose anal lobe ( Fig. 11 View Fig E–F), thinner paleae ( Figs 12A View Fig , 13A View Fig ) and the tip of the scaphal hooks is strongly curved (compare Figs 12F View Fig , 13F View Fig , 14F View Fig ).
The number and arrangement of teeth in the neuropodial uncini shows differences among BIOICE individuals, contrary to that observed in C. granulata . Two different types of uncini can be distinguished:
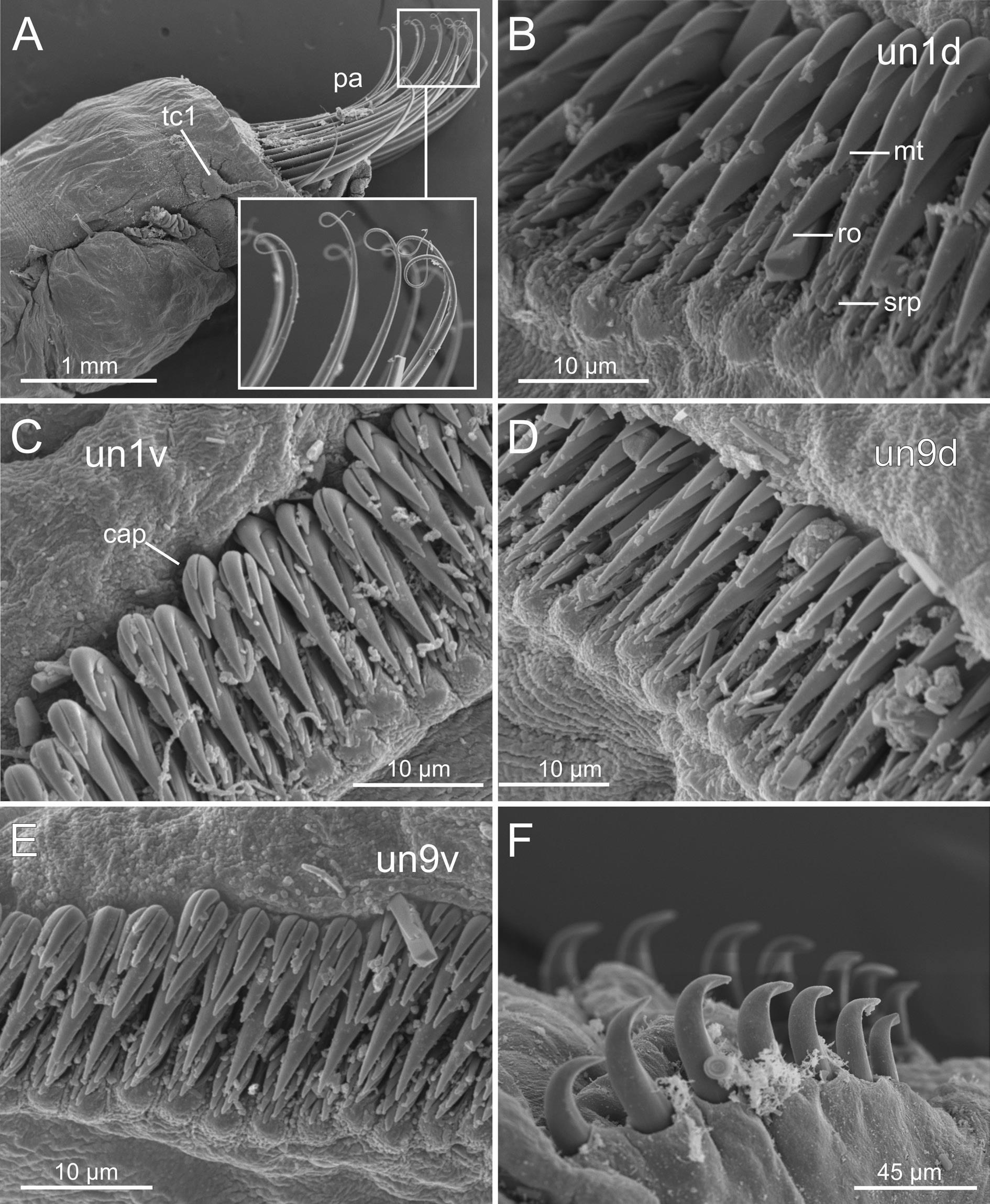
1) Uncini with 2–3 main teeth arranged in a vertical row. This is the typical teeth arrangement in the pectinate type for this species and the genus as well. We found this pattern in large specimens ( Figs 3D View Fig , 10 View Fig ).
2) Uncini provided with a large basal tooth (rostrum) and an upper group of short teeth (capitium). This agrees with the typical avicular type present in many species of Terebellomorpha and found here in smaller specimens ( Figs 3D View Fig , 12–13 View Fig View Fig ).
SEM examination of BIOICE specimens seems to support the hypothesis that these differences are related to body size even though we did not examine the whole range of sizes ( Fig. 3D View Fig ). Uncini of large specimens (sample 3252, Fig. 10 View Fig ) do not show a variation either within an individual torus or along the body; on the contrary, mid-sized specimens (sample 2660, Fig. 12 View Fig ) show a progressive increase in the number of upper teeth, mostly in ventral uncini ( Fig. 12C, E View Fig ) and especially in posterior chaetigers ( Fig. 12E View Fig ), the latter having a well-defined capitium; dorsal uncini show, in turn, the usual shape as found in the genus ( Fig. 12B, D View Fig ). This pattern of variation is more evident in small specimens (sample 2041, Fig. 13 View Fig ), where the dorsal uncini are also of avicular type ( Fig. 13 View Fig B–D), similar to ventral ones ( Fig. 13E View Fig ). Therefore, these results suggest that the BIOICE specimens of C. hyperborea show changes in the uncini shape during ontogenetic development and that the uncini type varies accordingly depending on the age of the specimen. This variation, as explained above, occurs within a torus and along the body; thus, as small specimens with the avicular type of uncini grow, and uncini are being replaced along the torus, there is a progressive decrease in the number of upper teeth and a loss of the rostrum finally resulting in a pectinate type with the typical formula of the genus (i.e., MF:1:1:1).
Specimens from sample 2060 ( Fig. 2D View Fig ) found in a shallow bottom ( 48 m) in North Iceland, were also identified as C. hyperborea . Nevertheless, they show thicker paleae (cf. Fig. 14 View Fig vs Fig. 13 View Fig ) than similarsized specimens from other samples, and dorsal uncini are similar to those of larger specimens found elsewhere ( Fig. 14B, D View Fig ), while ventral ones ( Fig. 14C, E View Fig ) correspond to those of specimens of their size such as those of sample 2660 ( Fig. 12C, E View Fig ). Finally, scaphe hooks appear more strongly curved than would be expected for this species (cf. Fig. 14F View Fig vs Figs 11E View Fig , 12F View Fig , 13F View Fig ). Therefore, this variation suggests that specimens from sample 2060 may correspond to another species (probably undescribed yet) within a potential species complex present in NE Atlantic waters, that should be assessed in future in combination with molecular studies.
Finally, SEM micrographs show that the tip of the notochaetae is serrated ( Fig. 15 View Fig A–B), and reveal the presence of perforations in the cuticle of the dorsal region anterior to the scaphe ( Fig. 15 View Fig C–D) and of two types of ciliated fields: 1) rounded fields located before the scaphe and among the cuticular perforations ( Fig. 15 View Fig C–D) and 2) one large field located dorsal to the anal lobe and anterior to the anal papilla ( Fig. 15 View Fig E–F). These fields may be related to water irrigation, but certainly a future in-depth study is necessary.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Cistenides hyperborea Malmgren, 1865
| Parapar, Julio, Palomanes, Verónica, Helgason, Gudmundur V. & Moreira, Juan 2020 |
Pectinaria ( Cistenides ) hyperborea
| Hartmann-Schroder G. 1996: 482 |
| Holthe T. 1986: 25 |
| Pettibone M. H. 1954: 312 |
| Nilsson D. 1928: 31 |
Pectinaria hyperborea
| Hessle C. 1917: 76 |
Cistenides hyperborea
| Malmgren A. J. 1865: 360 |