
Hebesuncus mollispinus, Pilato, Giovanni, Mcinnes, Sandra J. & Lisi, Oscar, 2012
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3446.1.4 |
|
DOI |
https://doi.org/10.5281/zenodo.5874140 |
|
persistent identifier |
https://treatment.plazi.org/id/03B887A6-FFD3-FFFE-B9BD-FC7CFD0DF816 |
|
treatment provided by |
Plazi |
|
scientific name |
Hebesuncus mollispinus |
| status |
sp. nov. |
Hebesuncus mollispinus sp. nov.
Material examined. Type locality: Vegetation: Moss – Coscinodon reflexidens Mull. Hal. ; Site: Unnamed outcrop in the Marion Nunataks, Charcot Island (69°45'S, 75°15'W), altitude: 150m. Collector: P. Convey, Date: 0 5 February 1999. Botanical sample in BAS Herbarium Database ( AAS Name and No.): Convey, P. 00326A.
Holotype: slide No. 5057, 8 paratypes (slide Nos. 5057 and 5058) and 16 eggs, 2 with the developed juveniles (slide No. 5058) deposited in the collection of Binda & Pilato (Museum of the Department of Animal Biology “Marcello La Greca”, University of Catania), a slide with three paratypes and one egg ( BASCH 012) deposited in the Natural History Museum, London (cat. No. BMNH ENT 2012-89), a slide with three paratypes and one egg ( BASCH 013) deposited in the National Museums of Scotland, Edinburgh (cat. No. NMS.Z.2011.160) and a further 41 paratypes and 10 eggs on 8 slides (slide Nos. BASCH 009-11; BASCH 019-22) deposited in the BAS Data Resource Collection.
Additional material: From mixed moss sample Charcot Island: Slide Nos. BASCH 030; BASCH 032; BASCH 034; BASCH 035. From Coscinodon sp. Alamode Island, Terra Firma Islands: Slide Nos. BASAL 001–2. From Andreaea sp. South Georgia: BASSG 0046.
Etymology. ‘mollispinus’ is derived from the Latin mollis – soft; spinus – spines, and refers to the soft spines observed on the eggs of this species.
Description of the holotype. Body length ca. 220 µm (not perfectly extended); colourless, eye spots present. Dorsal and lateral cuticle of the caudal region wrinkled with small, slightly protruding, irregular tubercles ( Fig. 2A View FIGURE 2. A – G ). This texture is more visible from legs III (legs II in one paratype) to the caudal extremity of the body. Buccopharyngeal apparatus is of the Hebesuncus type, i.e. with the bucco-pharyngeal tube subdivided into an anterior rigid portion (buccal tube) and a posterior, flexible, portion with a spiral thickening (pharyngeal tube) ( Fig. 2B View FIGURE 2. A – G ); buccal tube without ventral lamina and with the dorsal and ventral apophyses for the insertion of the stylet muscles as asymmetrical, blunt hooks, as described by Pilato (1987) ( Fig. 2B View FIGURE 2. A – G ). Drop-shaped thickening between the buccal and the pharyngeal tube absent; the narrowness of the tube obscures the junction between buccal tube and the start of the spirally thickened pharyngeal tube. The entire bucco-pharyngeal tube is 36.7 µm long and 1.3 µm in diameter (pbf = 3.5); with the stylet supports inserted at 46.4 % of the bucco-pharyngeal tube length. The pharyngeal bulb with well developed apophyses and two short macroplacoids; microplacoid and septulum absent. First macroplacoid 3.1 µm long (pbf = 8.4), second macroplacoid 2.3 µm long (pbf = 6.3); entire macroplacoid row 6.0 µm long (pbf = 16.3).
The claws, of the Hypsibius type, are small, with stout basal portion ( Fig.2D, E View FIGURE 2. A – G ) and main branches with well developed accessory points. External claws 8.3 µm long (pbf = 22.6) on legs II; 9.1 µm long (pbf = 24.8) on legs III. The main branch lengths of the external claws of legs I-III are 75% of the entire claw length (but difference in claw orientation gives paratypes values of 77.5–80.6%). Type specimen orientation prevents us from measuring inner claws and claws of leg IV, so we provide paratype measurements in Table 1 View TABLE 1 .
The eggs, freely laid, are spherical with flexible, thorn-like conical, and very rarely forked, processes ( Fig. 2F, G View FIGURE 2. A – G ) with diameters of ca. 62 µm excluding processes, and 75–80 µm inclusive. The processes (24–26 along the circumference and ca. 100 on the hemisphere) were 7.7–9.1 µm long with a basal diameter of 3.3–5.0 µm. Some processes have one, or more, transparent basal areas ( Fig. 2G View FIGURE 2. A – G ). The base of each process is indented ( Fig. 2G View FIGURE 2. A – G ) but process surface and the egg shell are smooth.
Remarks. The paratypes are similar to the holotype in both qualitative and metric characters. The measurements for some of the paratypes, including a juvenile, are given in Table 1 View TABLE 1 .
Differential diagnosis. Hebesuncus mollispinus sp. nov. differs from H. ryani in being colourless (H. ryani was described as light pink, or light violet to whitish), having a posterior wrinkled cuticle; bucco-pharyngeal tube slightly longer in relation to the body length ( Table 1 View TABLE 1 ); shorter and stouter claws ( Table 1 View TABLE 1 ) ( Figs.2D, E View FIGURE 2. A – G and 3B, C View FIGURE 3. A – F ); the external claws differ with the main branch inserted on the basal section at a lower position than those of H.
ryani ( Figs. 2E View FIGURE 2. A – G and 3B, C View FIGURE 3. A – F ). H. mollispinus sp. nov. also differs from H. ryani in egg shell details: the flexible, thorn-like processes are more numerous and uniform, wider basally and indented (diameter 3.3–5.0 µm in H. mollispinus sp. nov., 1.6–3.0 µm in H. ryani ) with fewer transparent areas (“bubble-like structures”— Dastych & Harris 1994), which are restricted to the basal portion of the process ( Figs. 2F, G View FIGURE 2. A – G and 3D–F View FIGURE 3. A – F ).
In his description of H. schusteri Dastych (1984) described the animal thus: “The body has light- or grey-pink colour, some specimens with lumps of brownish pigment.” There was no mention of cuticular ornamentation which we infer was absent. The poor condition of the studied paratypes did not allow us to discern cuticular ornamentation, so we cannot confirm the presence or absence of this character. Other robust characters however clearly differentiate this new taxon. Hebesuncus mollispinus sp. nov. differs from H. schusteri in that it is colourless and has a narrower bucco-pharyngeal tube (pbf values 3.4-3.9 versus 4.9 in H. schusteri ), shorter claws (the pbf values relative to the external claws on leg II are 22.6–25.2 in H. mollispinus sp. nov. and 35.9 in H. schusteri ; the pbf values relative to the posterior claws of legs IV are 30.4–30.8 in H. mollispinus sp. nov. and 48.6 in H. schusteri ). In addition, the eggs of H. mollispinus sp. nov. are completely different (the egg processes in H. mollispinus sp. nov. are conical, long and flexible while those of H. schusteri are hemispherical) ( Figs. 2F, G View FIGURE 2. A – G and 3 View FIGURE 3. A – F G).
There is some degree of confusion surrounding H. conjungens , which probably represents a species complex. We examined several specimens attributed to this species collected from Greenland and Italy (Apennines: Mount Rondinaio and Piedmont, Gran Paradiso: Colle del Nivolet), and we noted that the cuticle of these examples had a faint, roughness which is more or less evident under phase contrast ( Fig. 4A View FIGURE 4. A – D ). There is no mention of this character in the original description ( Thulin 1911), nor in any subsequent accounts of the species. It is not impossible that this was overlooked as it is sometimes very faint and would not be obvious under Khöler illumination. In the literature, most authors agree with Thulin’s description of the claws as, “kurz und dick” [small and stumpy] ( Thulin 1911) ( Fig. 4B, C View FIGURE 4. A – D , F). However, Maucci (1986), described Italian specimens thus: “il ramo principale è inserito piuttosto in alto, si che il ramo basale risulta piu lungo del secondario” [the main branches of the external claws are inserted aloft and, as a consequence, the basal tracts are longer than the secondary branches]. Horning et al. (1978) attributed a specimen from New Zealand to H. conjungens , with a body length of 235 µm and bucco-pharyngeal tube 54 µm long, whereas Dastych (1988) described specimens of H. conjungens from Poland having a similar body length (230 µm), but with the bucco-pharyngeal tube 36 µm long and 1.2 µm diameter. Our specimens from Greenland are 220 µm long, have bucco-pharyngeal tube 36 µm long and 1.7 µm diameter ( Fig. 4 View FIGURE 4. A – D E; Table 1 View TABLE 1 ). The eggs are usually defined as spherical, but can be oval (diameter 26-30 µm x 57 µm, according Cuénot (1932) and Maucci (1986)). The egg processes are generally defined as flexible spines, though Cunha (1941) described them as rigid on a specimen from Portugal. According to most authors the eggs of H. conjungens have thin, spine-like, well-spaced processes with basal diameter of ca. 2 µm (1.6–3 µm according to Dastych & Harris 1994). However, eggs we examined from Apennines have processes that are less well-spaced ( Fig. 4D View FIGURE 4. A – D ) than as they appear in the figures of many authors, for example Cuénot (1932, Fig. 73), Marcus (1936, Fig. 268D), Morgan & King (1976, Fig. 69), Dastych (1988, Fig. 108).
Notwithstanding this confusion, we can consider our specimens from Charcot Island different from H. conjungens in terms of egg characters. The eggs of H. mollispinus sp. nov. ( Fig. 2F, G View FIGURE 2. A – G ) have well developed, flexible, thorn-like processes with basal diameter of 3.3–5.5 µm, with transparent, “bubble-like” areas near the base and basal indentation; c.f. H. conjungens where these projections are spine-like, without transparent, “bubblelike” areas and more uniformly notched basal regions.
Conclusion. The addition of Hebesuncus mollispinus sp. nov. means that three of the four known Hebesuncus species ( H. schusteri , H. ryani and H. mollispinus sp. nov.) are confined to Antarctica. H. conjungens , traditionally considered as potentially cosmopolitan (with records from Europe, North America, Greenland, Hawaii, South America ( Argentina, Chile), New Zealand and [ Antarctica?]) ( Ramazzotti & Maucci, 1983; McInnes 1994)), has now a more restricted range. We concur with Dastych (1984, 1988), that the Southern Hemisphere records of H. conjungens need to be re-examined. Furthermore, we suggest that the Northern Hemisphere records of H. conjungens should also be revised, as the conflicting descriptions of this taxon appear to refer to more than one species.
Hebesuncus mollispinus sp. nov. is currently common to the southern maritime Antarctic and South Georgia (BAS Data Resource Collection). It is possible to speculate that the single specimens from King George Island ( Dastych 1984) and Joinville Island ( Usher and Dastych 1987) belong to this species but, without viewing specimens or eggs, this remains unconfirmed.
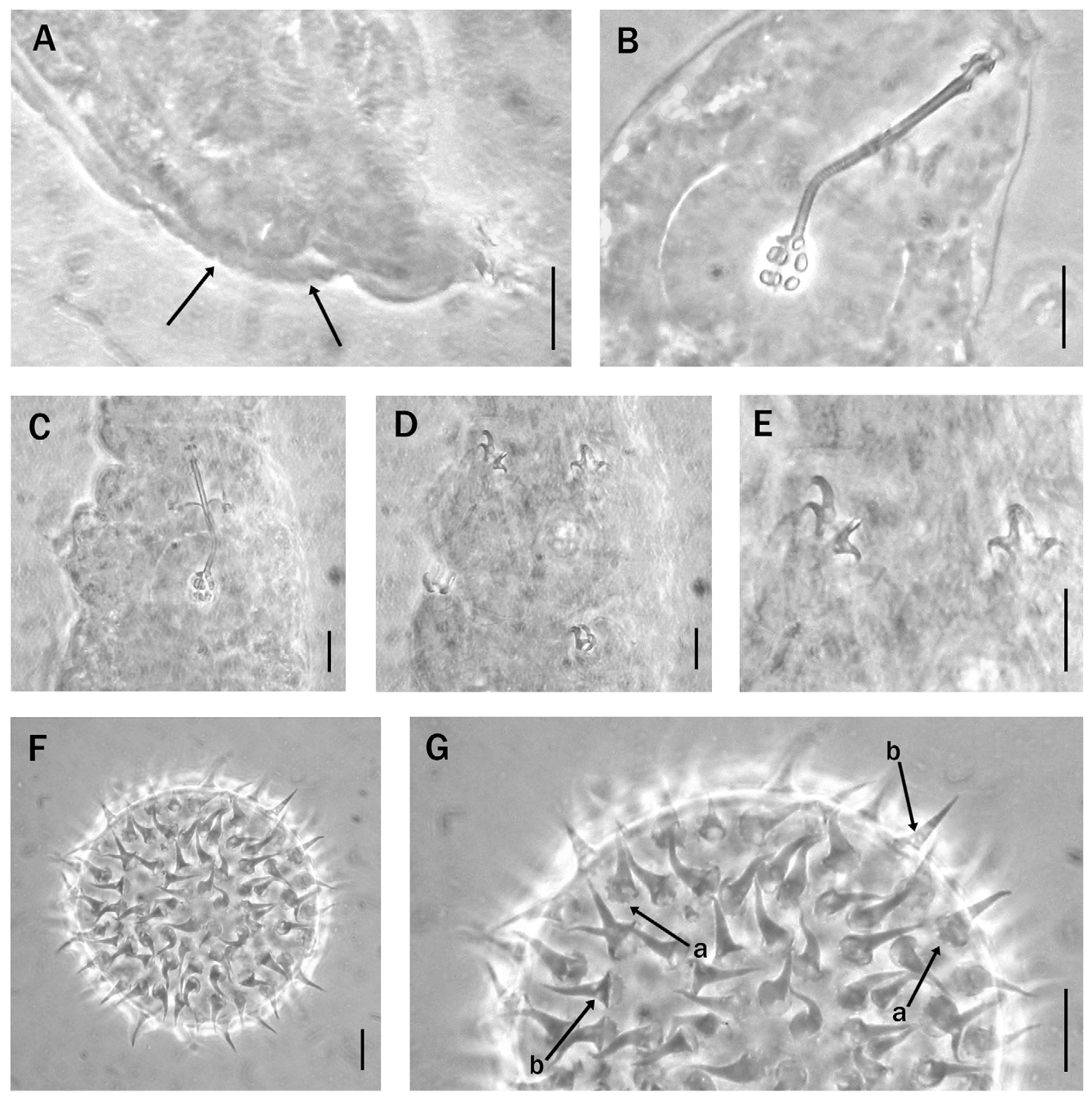
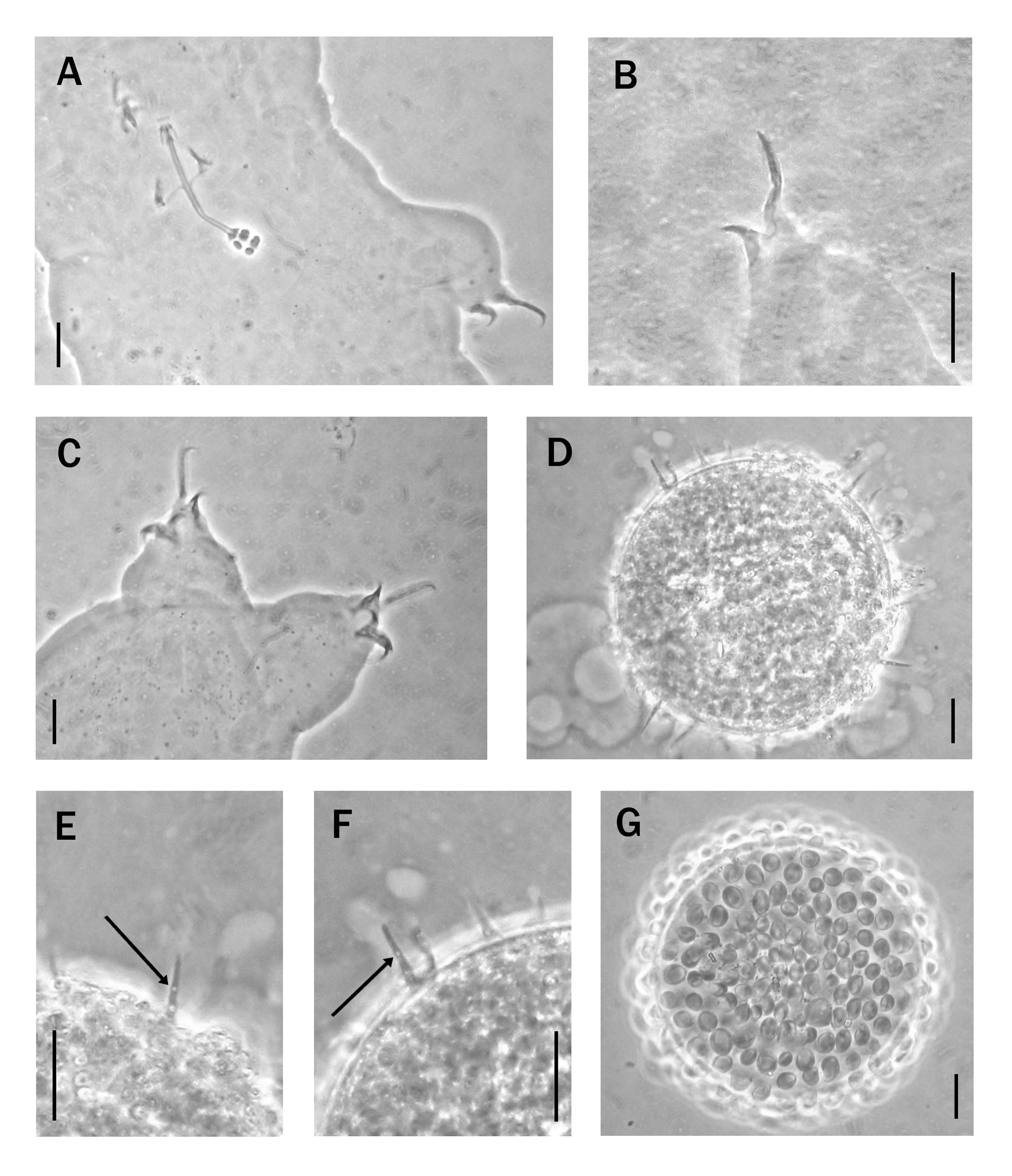

TABLE 1. Measurements (length in µm) of structures for Hebesuncus mollispinus sp. nov. and compared with Hebesuncus conjungens and Hebesuncus ryani of similar body length. pbf = The ratio between the length of a structure and the total length of the bucco-pharyngeal tube.? = orientation unsuitable for measurement
| Hebesuncus | |||||||
|---|---|---|---|---|---|---|---|
| mollispinus sp. nov. | conjungens Greenland | ryani Antarctica | |||||
| Slide No. | 5058 paratype juvenile | 5057 paratype | 5057 paratype | 5057 holotype | 1442 | 4301 paratype | 4301 paratype |
| Body length | 128 | ca. 180 | ca. 190 | ca. 220 | 220 | 230 | 261 |
| Bucco-pharyngeal tube length | 26.3 | 34.2 | 33.1 | 36.7 | 36.3 | 28.3 | 30.6 |
| Buccal tube width | 0.9 | 1.2 | 1.3 | 1.3 | 1.7 | 1.2 | 1.3 |
| pbf | 3.4 | 3.5 | 3.9 | 3.5 | 4.7 | 4.2 | 4.3 |
| Stylet supports pbf | 46.5 | 46.5 | 47.5 | 46.4 | 46.8 | 48.5 | 49.0 |
| Macroplacoids row | 4.0 | 5.7 | 5.8 | 6.0 | 6.5 | 5.0 | 5.6 |
| pft | 15.2 | 16.7 | 17.5 | 16.3 | 17.9 | 17.7 | 18.3 |
| First macroplacoid | 1.9 | 3.1 | 3.0 | 3.1 | 3.4 | 2.9 | 2.9 |
| pbf | 7.2 | 9.1 | 9.1 | 8.4 | 9.4 | 10.3 | 9.5 |
| Second macroplacoid | 1.7 | 2.0 | 2.5 | 2.3 | 2.7 | 2.2 | 2.4 |
| pbf | 6.5 | 5.9 | 7.6 | 6.3 | 7.4 | 7.8 | 7.8 |
| External claws II | 6.2 | 8.6 | 8.1 | 8.3 | 9.2 | ? | 14.5 |
| pbf | 23.6 | 25.2 | 24.5 | 22.6 | 25.3 | ? | 47.4 |
| Main branch | 5.0 | 6.7 | 6.4 | 6.2 | 7.1 | ? | 10.9 |
| % of total length | 80.6 | 77.9 | 79.0 | 75.0 | 77.2 | ? | 75.2 |
| Internal claw II | ? | ? | ? | ? | 6.6 | ? | 8.3 |
| pbf | ? | ? | ? | ? | 18.2 | ? | 27.1 |
| External claws III | 6.4 | ? | ? | 9.1 | ? | ? | 14.5 |
| pbf | 24.3 | ? | ? | 24.8 | ? | ? | 47.4 |
| Main branch | ? | ? | ? | 6.8 | ? | ? | 10.9 |
| % of total length | ? | ? | ? | 75.0 | ? | ? | 75.2 |
| Internal claws III | 4.1 | 6.1 | ? | ? | 7.0 | ? | 8.3 |
| pbf | 15.6 | 17.8 | ? | ? | 19.3 | ? | 27.1 |
| Posterior claws IV | ? | 10.4 | 10.2 | ? | ? | ? | 19.3 |
| pbf | ? | 30.4 | 30.8 | ? | ? | ? | 63.1 |
| Main branch | ? | ? | 7.3 | ? | ? | ? | 15.5 |
| % of total length | ? | ? | 77.5 | ? | ? | ? | 80.3 |
| Anterior claw | ? | 6.6 | 6.5 | ? | 6.7 | ? | 9.2 |
| pbf | ? | 19.3 | 19.6 | ? | 18.5 | ? | 30.1 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |