
Superodontella gladiator, Agolin, Mikaël, Houssin, Céline & Deharveng, Louis, 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.189772 |
|
DOI |
https://doi.org/10.5281/zenodo.6219894 |
|
persistent identifier |
https://treatment.plazi.org/id/03BB9D1C-FF80-9520-4AC1-FF2EFF46F819 |
|
treatment provided by |
Plazi |
|
scientific name |
Superodontella gladiator |
| status |
sp. nov. |
Superodontella gladiator sp. nov.
Figs 1–13 View FIGURES 1 – 9 View FIGURES 10 – 13
Type material. Holotype adult or subadult (with complete chaetotaxy but the presence of a genital plate cannot be confirmed due to the position of the furca), 4 adult or subadult paratypes mounted on the same slide as holotype; 2 paratypes prepared for SEM, 1 paratype in alcohol. Material deposited in the Muséum national d’Histoire naturelle de Paris.
Type locality. Nepal: Pulchauki, in Mahabarat range, near Godavari, alt. 2300 m., in thick humus and leaf litter of a Rhododendron forest, 26 x 1977 sample NEP77B-208, L. Deharveng leg.
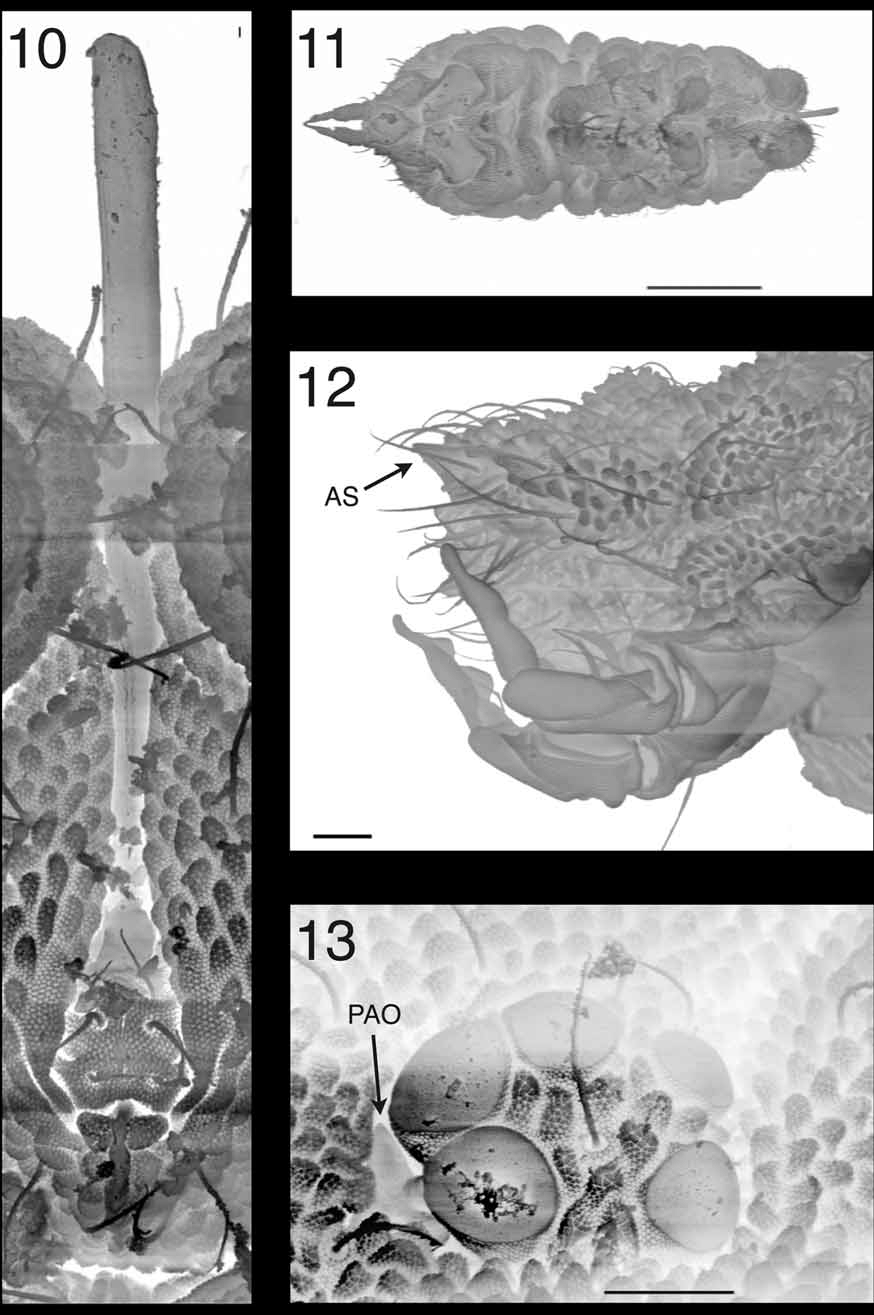
Description. Habitus typical of adult Superodontella ( Figs 1 View FIGURES 1 – 9 and 11 View FIGURES 10 – 13 ) as opposed to “large juvenile” form encountered in some species. Blue pigment sparse, more concentrated dorsally on head, Abd. V and VI, margins of other tergites—light blue color for animal in alcohol. Ventral side white. Ocular plates black.
Length of body (stylet excluded): 400-450 µm (specimens in scanning electron microscope), 700-800 µm (specimens on slides).
Integument with roundish granules, smallest on anterior part of head and gradually bigger and coarser to Abd. V and VI ( Fig. 12 View FIGURES 10 – 13 ). Slightly larger on central area of head and between Di and De areas of thoracic tergites ( Fig. 1 View FIGURES 1 – 9 ). A peculiar spindle-shaped patch of 10 to 15 very small granules (2 to 4 times smaller in diameter than the surrounding ones) at center of head, between eyes. Its length roughly equal to that of largest omma. Muscular insertions barely marked.
Antennae. Antenna IV dorso-distally with small ovoid organite (or) and a short microsensillum (ms) ( Fig. 3 View FIGURES 1 – 9 ); seven dorsal S-chaetae (S1, S2, S3, S4, S7, S8, S9 according to D’Haese 2003) rather long, bent, subequal, subcylindrical, and two ventro-basal chaetae akin to S-chaetae but less bent and more slender; a very long ventro-subapical blunt chaeta ( Figs 2 and 3 View FIGURES 1 – 9 ); 15-16 long, slender, more or less blunt dorsal mou-chaetae; 10- 11 short, pointed chaetae (including dorsal chaeta i), and 9 ventral, rather long, trumpet-chaetae. Antenna IV apex with a domed swelling of integument. Apical exertile vesicle absent. No eversible sac between segments III and IV. Sensory organ on antennal segment III with two short, bent sensory rods (S2 and S3), two short, bent, slightly longer and thinner guard sensillae (S1 and S4) and fifth dorso-external microsensillum (S5). S1, S2, S3 and S4 appressed to tegument in a slight depression. S1 bent externally, S2, S3 and S4 bent internally. Antennal segments I, II and III with respectively 7, 10 and 14 ordinary chaetae.
Head. Ocular plates with 5 omma each (A, B, C, F, G), with B the largest and G the smallest ( Figs 4 View FIGURES 1 – 9 and 13 View FIGURES 10 – 13 ). Each plate surrounded by a cuticular groove, circular in shape. Ocular grooves linked posteriorly by a transverse groove, anterior to c chaetae, posterior to d chaetae. Postantennal organ quadrilobed, roughly triangular owing to much smaller posterior lobe, sunk in a deep cavity in continuity with peri-ocular groove (visible on Fig. 13 View FIGURES 10 – 13 , bottom left corner), almost entirely covered: anteriorly by rim of cuticular invagination, posteriorly by overhanging anterior omma.
Mouthparts. Buccal cone styliform ( Figs 6 View FIGURES 1 – 9 and 10 View FIGURES 10 – 13 ): subcylindrical, slender and very long (as long as head). Very large clypeal area. Distal perilabial chaetae shifted closer to axis and rearranged around basis of buccal cone ( Fig. 6 View FIGURES 1 – 9 ). Perilabial chaetae b1 and b2 rather short, m1, b3 and b4 long and subequal. Postlabial chaeta m1 more than twice as long as b1. Chaeta p1 absent. Labium with only three small, subequal chaetae (probably E, F and G), and more distally with two small, spine-like structures. Whole labium elongated into two long, spoon-shaped lamellae forming a tube, slightly flattened dorso-ventrally at its tip. Mouthparts difficult to observe and recognize ( Fig. 5 View FIGURES 1 – 9 ). Two discernible parts: one clearly visible, sclerotized part, composed of styliform heads of maxillae, articulated to another, short and straight part (probably stipes) that is much less sclerotized and difficult to observe. Faint structures postero-internal to stipes (probably fulcra) and thin bilateral structures (maybe mandibles) antero-external to stipes. Mandibular head probably imbricated with maxillae head. Labrum very difficult to observe, with distal ridge but without visible chaetae.
Dorsal chaetotaxy as in Fig. 1 View FIGURES 1 – 9 . Ordinary chaetae short, subequal, smooth and pointed, longer and slightly serrated on abdomen VI. S-chaetae slightly longer, more refringent and twisted, 2,2/1,1,1,1,1 from thorax II to abdomen V ( Fig. 1 View FIGURES 1 – 9 ). Chaeta m1 present on thoracic tergites II, III and abdomen IV. On head, chaetae a0 absent, c2 and c3 present. Abdominal tergite VI plurichaetotic, with three rows of chaetae (one more than usual). Two anal spines ( Fig. 12 View FIGURES 10 – 13 ) short and pointed (probably modified granules).
Sternites and appendages. Leg chaetotaxy as in Table 1. Distal whorl of tibiotarsal chaetae ( Fig. 7 View FIGURES 1 – 9 ) with one acuminate tenent hair. Coxal chaetae not spiniform. Unguiculus absent. Claw with one ventro-basal tooth more or less appressed to internal crest, and two small latero-basal teeth. Ventral tube with three chaetae on each side. Sternite of abdomen II with (2) 3 chaetae Ve; of abdomen III with 5-6 chaetae Ve. Dens ( Fig. 8 View FIGURES 1 – 9 ) with 5 dorsal chaetae, three internal ones stronger. Mucro typical of the genus with its two intermediate lamellae, and as long as dens. Chaetotaxy of anal valves and ventral abdomen VI as in Fig. 9 View FIGURES 1 – 9 . Lateral valves with three hr chaetae, dorsal valve with only two observable hr chaetae.
Etymology. This species is named because of its long, thin, slightly flattened labial sheath, reminiscent of the spatha, a straight sword with a long point wielded by the ancient gladiators.
Discussion. The species described here is typical of the genus Superodontella , but at the same time quite peculiar. It bears all the diagnostic characters of the genus: 5+5 pigmented ommatidia, no eversible sac between Ant. III and IV, no apical vesicle on Ant. IV, strongly modified labium and mouthparts, 4-lobed postantennal organ, tibiotarsus with two whorls of chaetae (as in all other non-regressed genera), a well developed furca, dens with 5 dorsal chaetae, and a large mucro with two dorsal lobes. It belongs to Deharveng’s (1981a) group 3, with two anal spines, three of the five dental chaetae thickened and an elongated buccal cone. Furthermore, the species for which Stach (1949) erected the genus Superodontella , Odontella ewingi Folsom, 1916 from the USA, also pertains to group 3.
It is, however, peculiar, because it is the first described Superodontella to exhibit such a level of mouthpart elongation, stipes modification and three rows of chaetae on the tergite of Abd. VI (although Odontella setosa Deharveng, 1981 b also has a plurichaetotic Abd. VI). An unidentified and undescribed species from Singapore characterized with the mouthparts similar to S. gladiator sp. nov. is studied in an unpublished Bachelor of Science report by Sim Jui Eng (1970).
Table 2 gives a comparison of S. gladiator sp. nov. with the other Asiatic Superodontella sporting very long beaks, regressed labial chaetotaxy, short anal spines and an apical swelling of the integument of Ant. IV: S. similis Yosii, 1954 from Japan, S. cornuta Yosii, 1965 from Japan, S. nepalica Yosii, 1971 from Nepal, and S. ciconia Bedos & Deharveng, 1990 from Thailand.
The type of mouthparts described herein, for which the term “ gladiator mouthparts” is coined, differs from the "superodontellian" type of Deharveng (1981a) and can be defined as follows: stipes and fulcrum well developed, clearly discernible and directly articulated to each other as in all other Odontellidae , sometimes lacking one of the two mandibles, and with the fulcrum postero-internal process much longer than its posteroexternal process.
For the moment, without a dissection of the gladiator mouthparts, their organization is unclear. What is evident, however, is the presence of an unusual articulation between stipes and maxilla head ( Fig. 5 View FIGURES 1 – 9 ), not recorded in any other Poduromorpha . This provides support for the interesting hypothesis of Sim Jui Eng (1970) who, following the dissection of the mouthparts of Superodontella spp., proposed that the stylet itself is made up of the stipes of the maxilla, the finely striated and bevelled extremity being its head. In that case, what we describe as short, sclerotized, straight stipes could in fact be the cardo. However, the original drawing of Deharveng (1981a, Fig. 1 View FIGURES 1 – 9 ) for fresh Nepalese specimens of S. gladiator shows an armature of maxillae not very different from that of other species of the genus. We were unable to discern the structures described 27 years ago, which may be due to deterioration of the preparation, or incorrect observation. The examination and dissection of fresh specimens of S. gladiator will be necessary to clarify this point.
S. gladiator S. ciconia S. cornuta S. nepalica S. similis Spines on Ant. I and no no yes? no II
Dorsal S-chaetae on 7 7 ca. 10? 5-6 Ant. IV *
Type of mouthparts gladiator superodontellian?? superodontellian Rows of chaetae on 3 3 3 2 3 Abd. IV
* The number of S-chaetae on Ant. IV given in the literature for Superodontella species should be taken with caution, as explained in the text for S. gladiator .
Another characteristic described in Sim Jui Eng’s work and apparently shared by S. gladiator is the close imbrication of mandibles, fulcra and cardos (or stipes?) at the base of the buccal cone, from where the maxillae and mandibles elongate into the labial sheath. Unfortunately, we were unable to follow the projection of the faint mandibles in this sheath, if present, even at high magnification.
Our observations on the structure of the labium on the slide were confirmed by scanning electron microscope, especially its three-dimensional shape and the rotation of the two hemilabia. The three chaetae are clearly visible. We hypothesize that they are E, F and G because in other Superodontella with an elongated beak, they are almost always present, and the last to regress. However, on such a modified labium, there is no way we could give them a definite name. The more distal spine-like structures on the labium could not be observed by SEM, probably because they are internal. A remarkable feature of the S. gladiator mouthparts is that they emerge from the head very ventrally, almost at its center. However, the stylet is still oriented straight ahead, owing to the presence of a groove in the clypeus into which it is sunk, as shown in Fig. 10 View FIGURES 10 – 13 . Such a structure raises many questions concerning its function. Can it be orientated? Is this stylet used to pierce and suck, and if so, what? In fact, shorter mouthparts in other Superodontella species also exhibit a posterior shift, but they emergence from the head less posteriorly––a character that was overlooked in previous studies. Such unusual morphological evolutions probably have a function in the feeding habits of the species concerned, which remains to be elucidated.
We have not named the three ventral S-chaetae of the fourth antennal segment because they do not fit into D’Haese’s (2003) unified system for chaetotaxic homologies in Poduromorpha . In S. gladiator , as in many other Superodontella species, some chaetae on Ant. IV tend to be longer, blunt and more refringent than ordinary chaetae, looking like thin S-chaetae. Intermediate stages may be observed, making it sometimes difficult to distinguish them from true S-chaetae. These chaetae include those named mou-chaetae by Deharveng (1983) in Neanurinae and the conspicuous, long, subapical macrochaeta present in several Superodontella species and other Poduromopha. In contrast, ordinary chaetae are consistently shorter and pointed.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |