
Guestphalinus exilis Fend & Rodriguez, 2017
|
publication ID |
https://doi.org/10.5852/ejt.2017.361 |
|
publication LSID |
lsid:zoobank.org:pub:F61CB5C7-B22E-4FAB-997A-BF99C7828C77 |
|
DOI |
https://doi.org/10.5281/zenodo.3851836 |
|
persistent identifier |
https://treatment.plazi.org/id/37A5A3E0-3DC2-4D24-84AC-62E21131E70E |
|
taxon LSID |
lsid:zoobank.org:act:37A5A3E0-3DC2-4D24-84AC-62E21131E70E |
|
treatment provided by |
Carolina |
|
scientific name |
Guestphalinus exilis Fend & Rodriguez |
| status |
sp. nov. |
Guestphalinus exilis Fend & Rodriguez View in CoL sp. nov.
urn:lsid:zoobank.org:act:
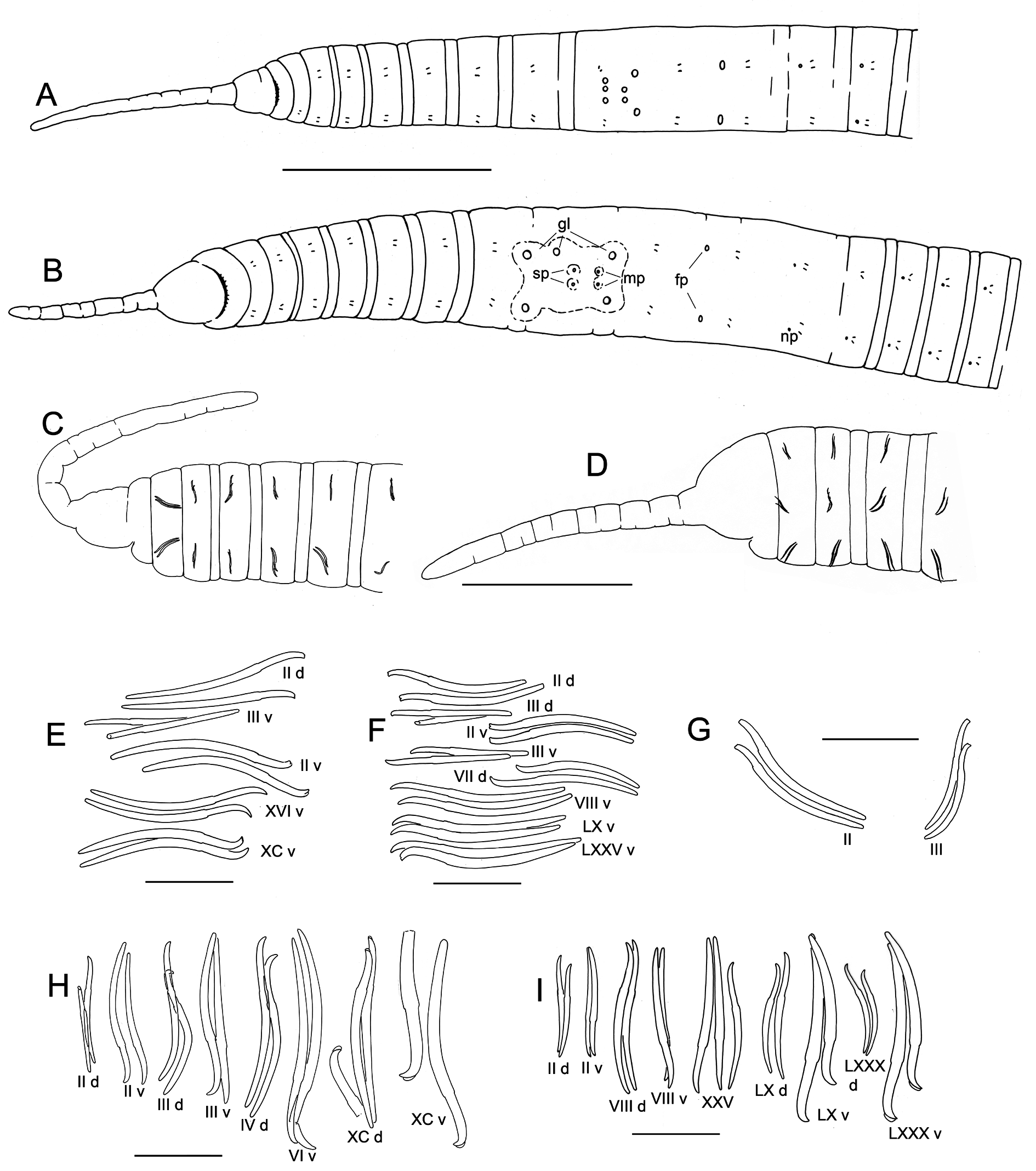
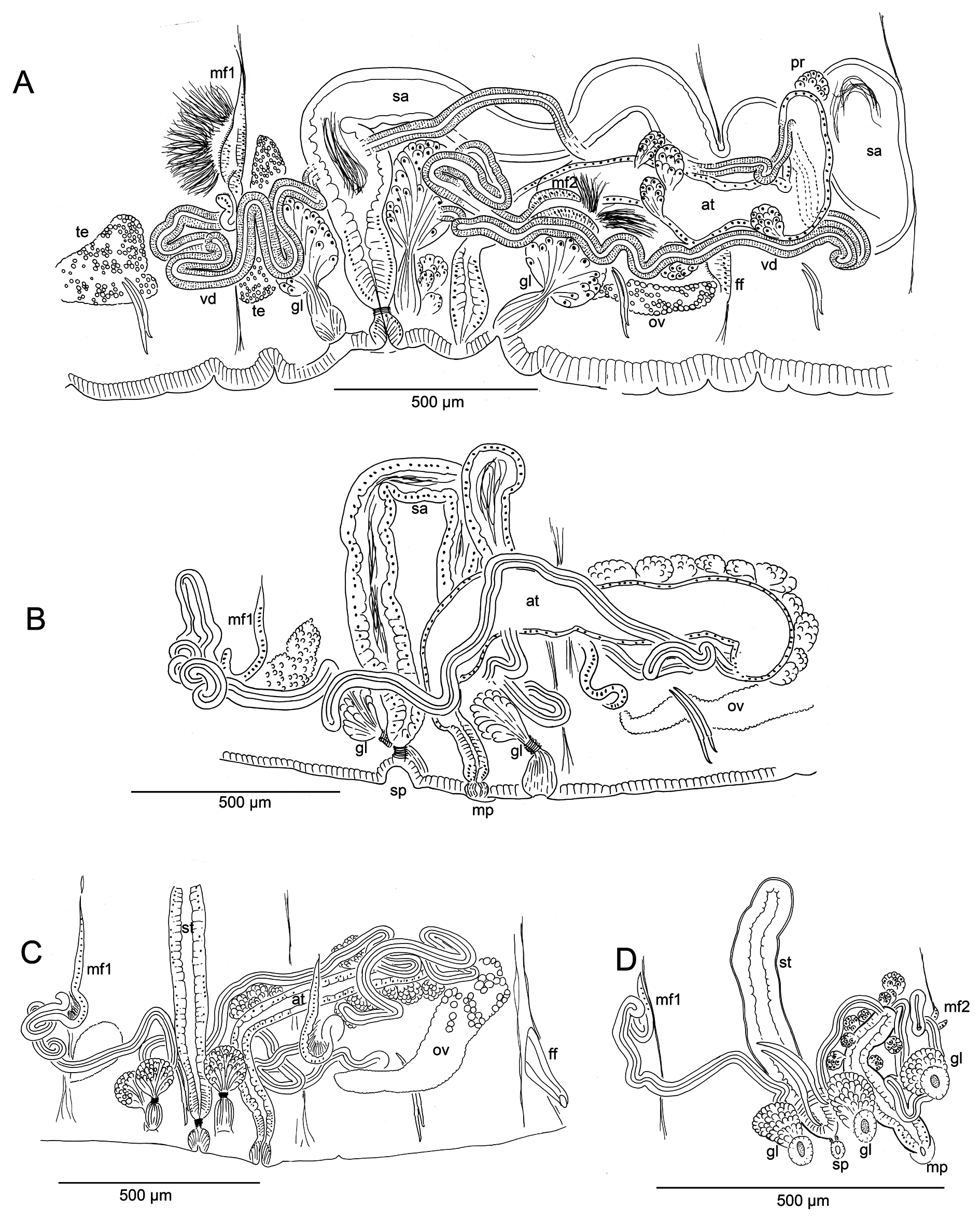
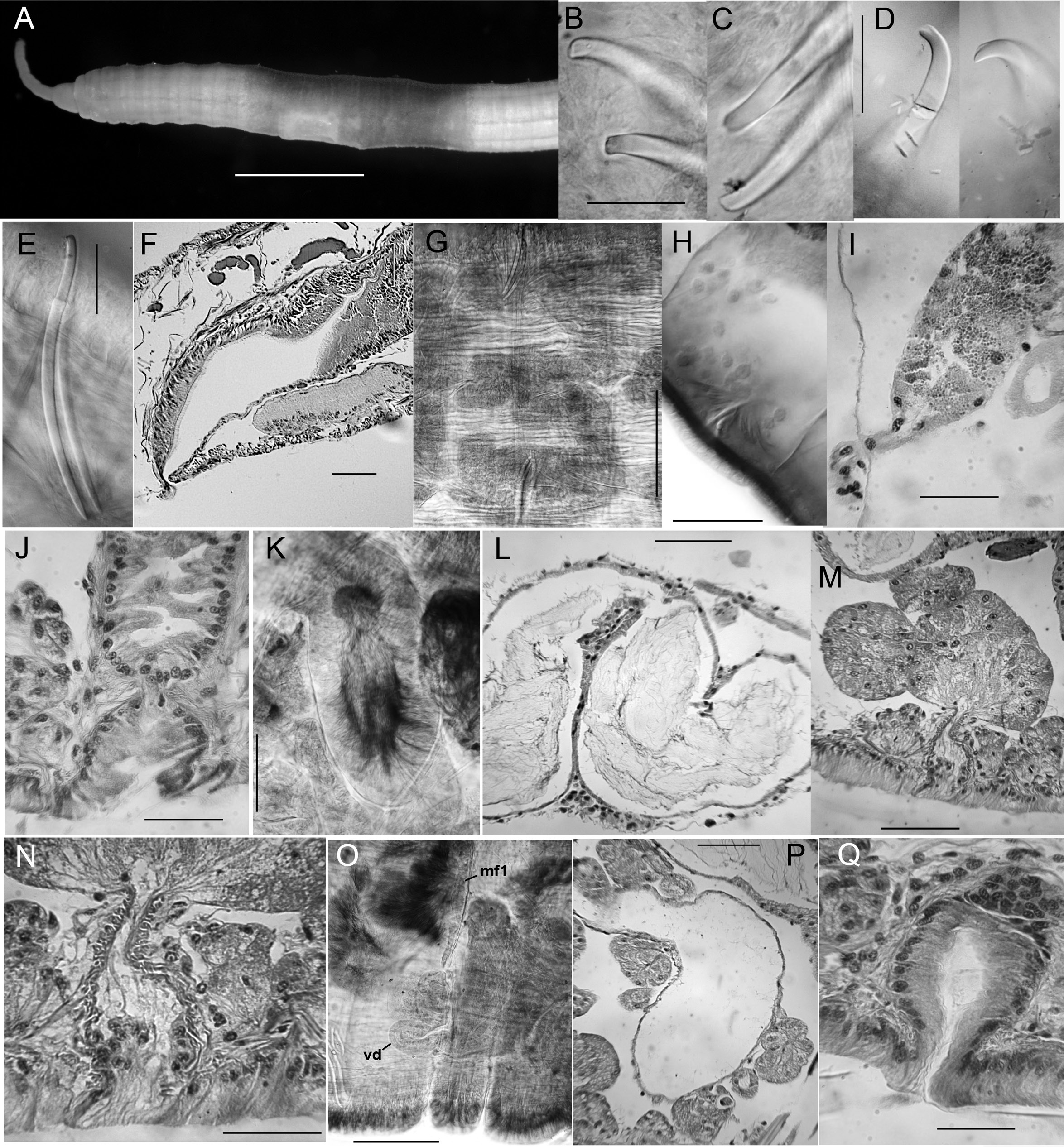
Figs 7–9 View Fig View Fig View Fig
Etymology
Derived fom the Latin word ‘ exilis ’ (small), with reference to G. elephantinus sp. nov. (see below) and to the name of the extinct Californian pygmy mammoth Mammuthus exilis (Stock & Furlong, 1928) .
Material examined
Holotype
UNITED STATES OF AMERICA: a dissected specimen, slide-mounted in Canada balsam, California, Shasta County, Squaw Creek at Chirpchatter Camp , deep riffle, gravel patches in cobble bottom, 6 May 2012, S. Fend leg. ( USNM 1422283 About USNM ).
Paratypes (all collected by S. Fend at the type locality)
UNITED STATES OF AMERICA: 1 dissected on 2 slides, 19 Apr. 2010 ( USNM 1422284); 1 dissected, 17 May 2010 ( USNM 1422285); 1 dissected, 6 May 2012 ( CASIZ 220935); 1 whole mount, 17 May 2010 ( CASIZ 220936); 1 sagittally sectioned, 17 May 2010 ( CASIZ 220938); a whole mount, immature but with gonads (DNA voucher), 19 Apr. 2010 ( CASIZ 220937); 2 dissected on slides, 17 May 2010 ( MNCN 16.03/3100–16.03/3101).
Additional material
UNITED STATES OF AMERICA: CALIFORNIA: 2 sagittal sections, 3 dissected, 4 whole mounts (all immature or partially mature), Shasta County, Squaw Creek at Chirpchatter Camp, 17 Jun. 1996, S. Fend leg.; 1 dissected (partially mature), same locality as preceding, 19 Apr. 2010, S. Fend leg.; 4 dissected (1 mature), 8 whole mounts (immature), several in alcohol, same locality as preceding, 17 May 2010, S. Fend leg.; 1 dissected (mature), several immature in alcohol, same locality as preceding, 6 May 2012, S. Fend leg.; 2 whole mounts (partially mature), Humboldt County, South Fork Eel River at Elk Creek, 24 Jun. 2001, S. Fend leg.; 1 whole mount (partially mature), Mendocino County, Garcia River, 23 Mar. 1996, W. Fields leg.; 1 whole mount (immature, with small gonads), Colusa County, Stony Creek at Stonyford, 4 Nov. 2002, S. Fend leg.
Molecular data
COI, 28S and 16S sequences correspond to a topotypic paratype (see Table 1 View Table 1 for details).
Description
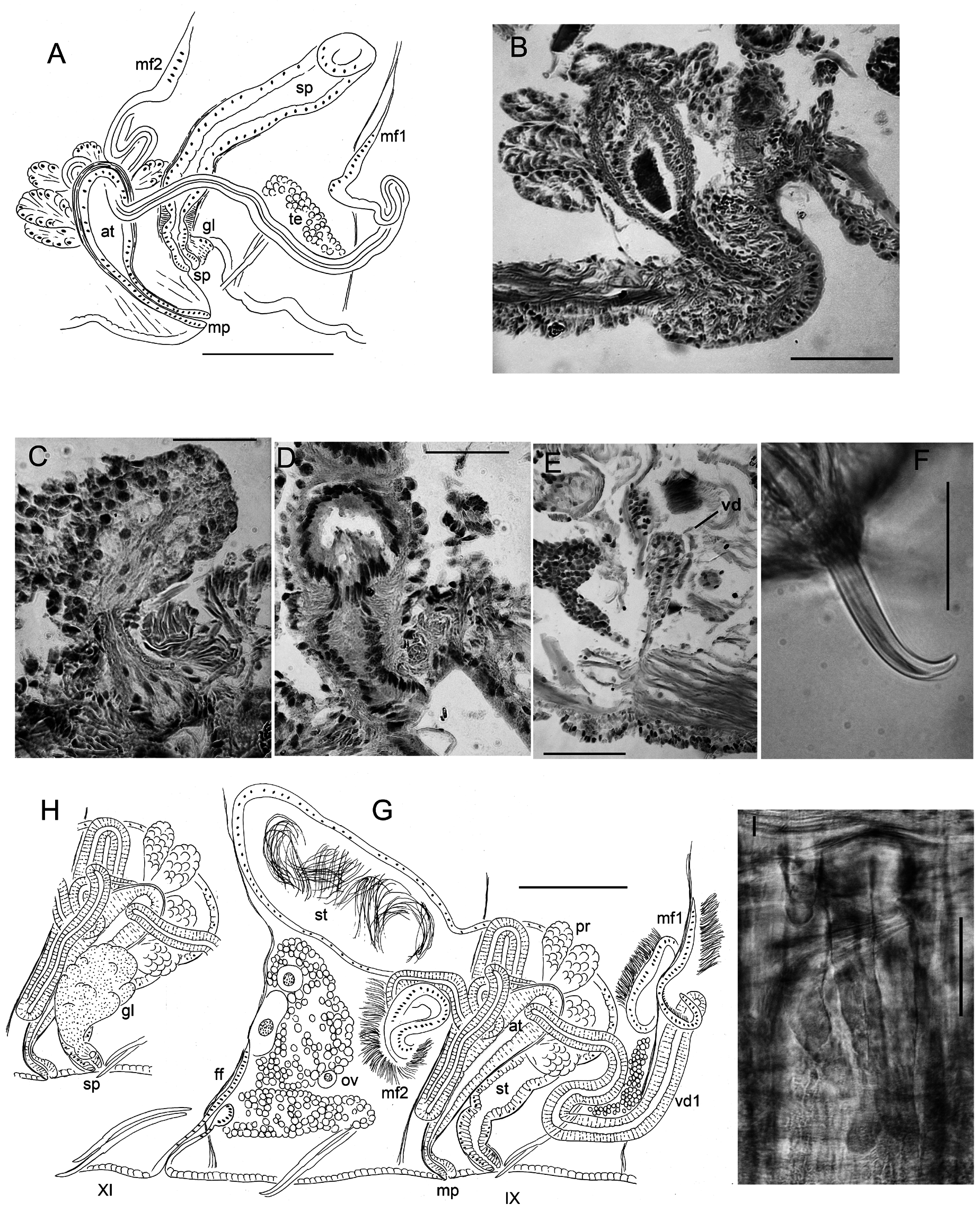
Specimens from the type locality: body measurements in Table 3. Prostomium rounded to nearly conical; filiform proboscis 0.9 to 2.1 mm long, diameter at midpoint 0.1–0.15 mm, appearing ringed with multiple shallow constrictions ( Figs 7 View Fig A–C, 9A). Body segmentation not strong in external view; secondary annulation a narrow anterior ring in IV–IX, weak in post-clitellar segments. Clitellum from VIII to XII or mid-XIII, absent ventrally in IX in the area surrounding male and spermathecal pores ( Figs 7B View Fig , 9A View Fig ).
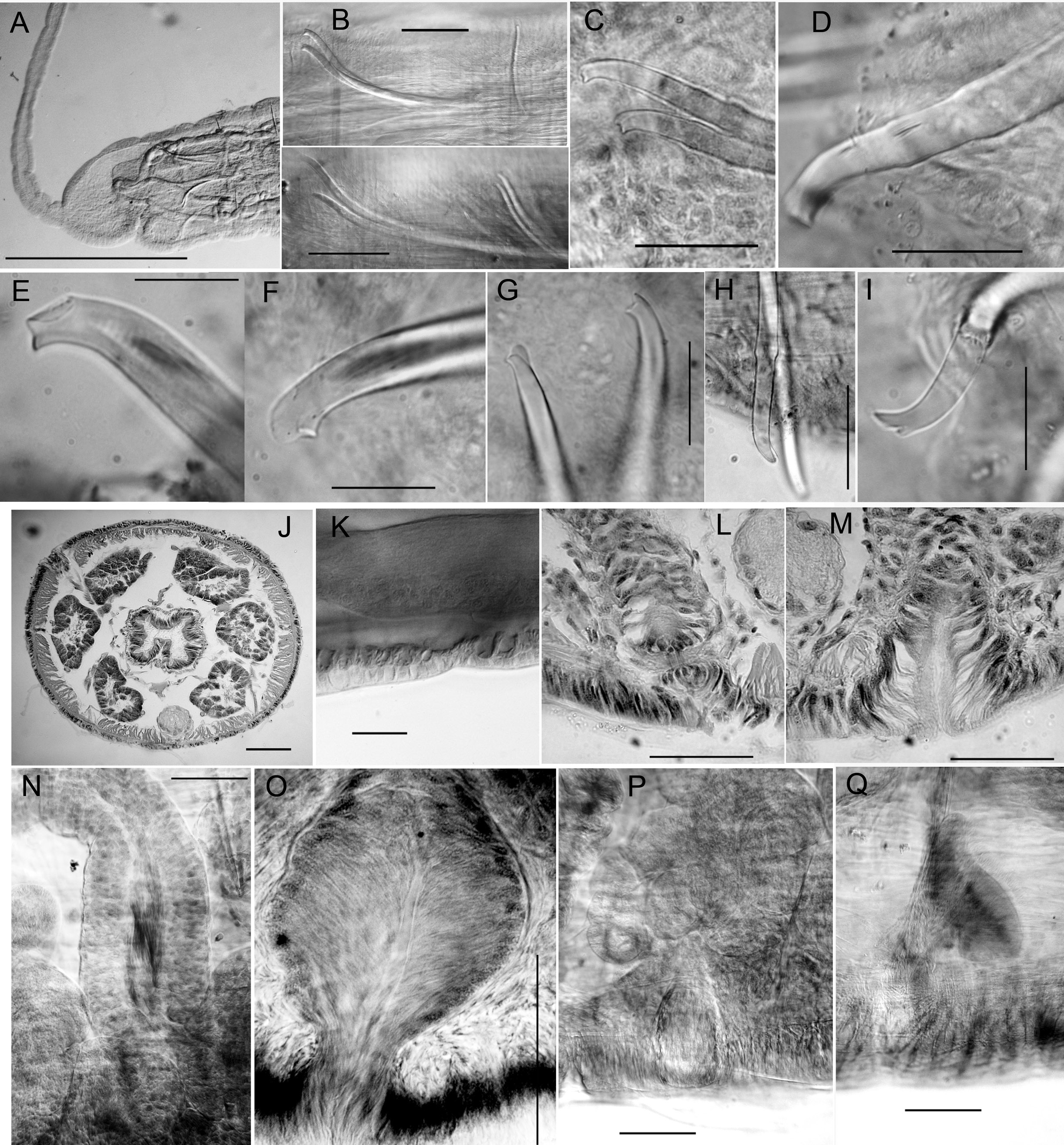
Chaetae paired, in 4 bundles in each segment, beginning in II. Chaetal measurements given in Table 3. Chaetae in II bluntly simple-pointed to shallowly notched, directed anteriorly ( Figs 7 View Fig E–G, 9B–C, E); these chaetae slightly sigmoid, with distal nodulus; within each bundle, lateral chaeta slightly longer than the median, with more distal nodulus. Posterior to II, chaetae sigmoid, mostly simple pointed, with nodulus about ⅓ of distance from tip; perpendicular to body axis or posteriorly directed; chaetae in III slightly shorter, but those in more posterior segments similar in length to those in II. Chaetae in posterior segments may have a slight dorsal keel ( Fig. 9D View Fig ). Ventral chaetae absent in IX in mature and post-reproductive worms.
Epidermis in anterior segments 12–24 µm thick, in clitellum 30–50 µm, posteriorly 5–10 µm. In preclitellar segments, circular muscle layer of body wall arranged in a series of transverse bands (cf. Fig. 12K View Fig ), 10–15 µm thick; posteriorly a simple layer about 5 μm thick. Longitudinal muscle layer 50–60 µm thick. Brain in the peristomium, lateral lobes rounded. Pharynx begins dorsally and laterally in II, ventrally in III, extending through IV. Pharyngeal glands in IV to VI or VII; on each side, three lobes (dorsal, lateral and median) ( Fig. 9G View Fig ) broadly connected at posterior septum of each segment and extending anteriorly, joining corresponding lobe in previous segment by a thin extension. No abrupt division between esophagus and intestine. Chloragogen cells cover the gut usually beginning in VII; in the most posterior segments many free eleocytes present in the coelomic cavity ( Fig. 9H View Fig ).
First nephridia usually on 6/7, absent in VIII–XI, usually paired on 11/12, occuring irregularly in posterior segments. Each nephridium with small anteseptal funnel; granular postseptal expansion elongate to ovate (length 130–250 µm, diameter 40–80 μm), directed posteriad or somewhat dorsad ( Fig. 9I View Fig ); convoluted efferent duct may pass through one or more adjacent (anterior or posterior) segments, ventral or ventrolateral to the gut, usually near the ventral blood vessel, terminating in a short ectal branch to a simple nephridiopore anterior to the ventral chaetae; indistinct vesicle at the pore in some specimens.
Dorsal blood vessel passes under brain: two forks pass around the pharynx and join in IV, forming the ventral vessel. Dorsal vessel free anteriorly, closely appressed to top of gut posterior to VI or VII. Ventral vessel separate from gut, but 2–3 short vessels join it to the perivisceral sinus in each segment posterior to about VII. One pair of commissural vessels in anterior segments; those in II–VI (or VII) long and sinuous, extending through most of originating segment; those in VIII to XII (or XIV) restricted to posterior part of the segment, but a posterior loop from those in IX and X may enter sperm and egg sacs. No lateral blood vessels observed behind about segment XIV. Perivisceral sinus begins in about VII.
Genital field covers ventral side of IX, flattened or slightly concave in preserved, mature worms. Male and spermathecal pores paired in IX. Male pores near posterior intersegmental groove ( Fig. 7 View Fig A–B) and inside lines of ventral chaetae; spermathecal pores in line with, and in front of male pores, about level with dorsal chaetal bundles (ventral chaetae absent in IX). Female pores paired, on chaetal line at intersegmental groove 10/11. Secretory openings of copulatory glands in IX, small and circular areas, lateral to genital pores, and either in front of or behind them. Paired testes on anterior septa in VIII and IX may be large, extending through anterior half of segment; ovaries in X, reaching to posterior septum, or into XI. Sperm sacs extending back to XIV–XVII in mature worms, egg sacs as far as XV–XVII.
Spermathecae to over 2000 µm long in mated worms, nearly tubular, with two, weakly differentiated sections, both containing sperm. Ectal, duct-like section gradually expanding to form ental ampulla; ampulla may be folded within IX (both in unmated or mated specimens) or extending into X–XII. Ectally, spermatheca passes through a short (12 μm), narrow constriction within ring of muscle fibers ( Figs 8 View Fig A–B, 9J) and terminates in a shallow epidermal infolding, about 70–110 μm deep. Duct-like section (80–100 μm diameter) with irregular, columnar epithelium (up to 40–45 μm high); dense sperm may be lined up along epithelium ( Fig. 9K View Fig ); very thin outer muscle layer about 2 μm thick. Ampullalike section nearly tubular, diameter about 100 µm for much of its length, but may be entally expanded to over 200 μm ( Figs 8A View Fig , 9L View Fig ), with thinner epithelium and wider lumen containing sparse spermatozoa.
Typically, 3–6 stalked copulatory glands are associated with genital pores in mature and nearly-mature worms ( Fig. 8 View Fig A–C); they are absent at early stages of reproductive development. Glands are a pyriform cluster of granular, petiolate cells 160–300 μm long ( Fig. 9 View Fig M–N), sometimes appearing as a group of smaller clusters. Conjoined cell extensions of the gland are constricted by a ring of circular muscle fibers and then surrounded by thin epithelium before opening in a round secretory surface (to 50 µm diameter) on the body wall.
Anterior and posterior male funnels about equal in size, to 240 μm high; anterior pair rather flat on septum 8/9; posterior pair on 9/10, but usually directed back into X; both pairs functional, with sperm when fully mature. Anterior vas deferens extends into VIII ( Figs 8 View Fig A–C, 9O), forming a convoluted mass, then penetrates 8/9, running along ventral body wall to near the male pore, then follows atrium to near the ental end, joining the atrium apically. Length of anterior vasa deferentia to 2600 μm, width 36–50 μm. Posterior vas deferens forms a compact, convoluted mass in posterior IX, then follows atrium within the sperm sac, joining it at or near the apical end. Length of posterior vasa over 2000 μm, width 38–46 μm. Vasa deferentia histologically similar throughout, with ciliated, non-glandular epithelium.
Atria of mature worms usually extend back into X or XI; in nearly-mature worms they may be entirely in IX. Atria petiolate in mature worms ( Fig. 8 View Fig A–B); a short ectal duct (150 μm long) has thick, columnar epithelium and a thin (to 2 μm) muscle coat; no distinct penis, although duct may be somewhat expanded at male pore, with thickened epithelium ( Figs 8B View Fig , 9Q View Fig ). An abrupt transition from the atrial duct to the tubular or sacciform atrial ampulla. Ampulla length to 1060–1105 μm, width 130–150 μm, thin-walled (5–8 μm) with cuboidal epithelium and wide lumen ( Figs 8A View Fig , 9P View Fig ); clusters of prostatic cells, 70–140 μm long, sparsely cover the atrial ampulla. In nearly-mature, unmated worms, atria tubular ( Fig. 8C View Fig ); ampulla and duct not greatly differentiated; prostates small but appear more densely packed than in mated worms.
Eel River, Stony Creek and Garcia River, northern California: The few, partially-mature specimens from these localities are tentatively assigned to Guestphalinus exilis sp. nov. based on the ringed proboscis, the chaetae, and gonads in VIII, IX and X. They differ from the type diagnosis in that chaetae in II are smaller than other anterior chaetae, and simple-pointed (but oriented forward, as in the type locality population). Ventral chaetae in posterior segments distinctly keeled and larger than corresponding dorsals ( Fig. 7 View Fig H–I). As in typical G. exilis sp. nov., specimens from Eel River have semiprosoporous male ducts, with vasa deferentia joining atrium apically and the anterior vas deferens penetrating 8/9 ( Fig. 8D View Fig ). Spermathecal and male pores are behind ventral chaetae of IX and 5 large copulatory glands surround the genital pores. One specimen has an additional, developing spermatheca on one side, in X.
Remarks
Guestphalinus exilis sp. nov. from western North America resembles the Palearctic G. wiardi in characters considered diagnostic for the genus (see above). Compared with G. wiardi , G. exilis sp. nov. has modified chaetae in II, a much larger, sacciform atrium, and more copulatory glands; the male pore was never strongly protruding ( Fig. 6B View Fig vs Fig. 8 View Fig A–B). The reproductive organs of G. exilis sp. nov. are morphologically similar to those of G. elephantinus sp. nov. (see below); both lack a distinct male porophore or penis, although the atrial duct may be somewhat expanded at the male pore, with thickened epithelium, suggesting that lining cells may be protrusible ( Figs 9Q View Fig , 12M, O View Fig ) as “type-2 penes” (temporary structures formed by extruded lining cells of the male duct, see Rodriguez & Giani 1994: fig. 17D–F).
Molecular results strongly suggest more than one Nearctic species (see below). Both new Guestphalinus species present distinctive chaetal morphotypes, and these chaetal characters are therefore used here as main diagnostic characters. Nevertheless, there is some variation among populations assigned to both basic morphotypes. In segment II chaetae of typical (from Squaw Creek) G. exilis sp. nov. are slightly modified relative to those in other segments. In most Squaw Creek specimens the chaetae appear slightly notched, slightly larger than those in III, and a little shorter than those in middle segments ( Table 3). In specimens from two other California sites, chaetae in II are simple-pointed and smaller than those in III – and thus similar to most other lumbriculids. These are provisionally assigned to G. exilis sp. nov., based on the limited and only partially-mature available material. Partially developed reproductive organs ( Fig. 8D View Fig ) clearly indicate that the Eel River specimens belong to Guestphalinus as defined here. The single specimens from Stony Creek and the Garcia River, although immature, have gonads in VIII–X, distinguishing them from other proboscis-bearing lumbriculid genera in the region (e.g., Kincaidiana , Rhynchelmis and Eremidrilus ).
Habitat
The type locality is Squaw Creek, a tributary to the Sacramento River, in northern California; the collection site is a 5 th order riffle-pool stream with gravel-cobble sediment ( U.S. D.A. 1999). The type series was found only at the lower end of a single riffle, and no additional material has been found at other sites within the stream. Other oligochaetes were collected throughout the stream; dominant species were Mesenchytraeus pedatus Eisen, 1904 and Rhyacodrilus clio Rodriguez & Fend, 2013 (also the type locality for R. clio ). The four streams where G. exilis sp. nov. was collected were similar in size and sediment characteristics, and are coldwater streams, supporting salmonid fishes. The species was usually found in patches of finer gravel within cobble-boulder riffles. Specific conductance at the Squaw Creek and Eel River sites ( April 2014) was 265 and 250 µS cm-1, respectively. Earlier measurements at a downstream site on Squaw Creek ( Rettig & Bortleson 1983) gave temperatures from 13.5 to 22.5 °C and a specific conductance of 109–261 µS cm-1. Visits to the type locality from July to September did not produce any specimens of G. exilis sp. nov. The sporadic occurrence and patchy distribution suggest that, like G. wiardi , they may also be primarily groundwater or hyporheic worms that only occasionally show up in surface collections.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |