
Ctenomys perrensi, Thomas, 1896
|
publication ID |
https://doi.org/10.1515/mammalia-2016-0162 |
|
DOI |
https://doi.org/10.5281/zenodo.7859435 |
|
persistent identifier |
https://treatment.plazi.org/id/03BC87CB-C063-FFB9-229A-EA867648FC9E |
|
treatment provided by |
Felipe |
|
scientific name |
Ctenomys perrensi |
| status |
|
Ctenomys perrensi View in CoL View at ENA complex
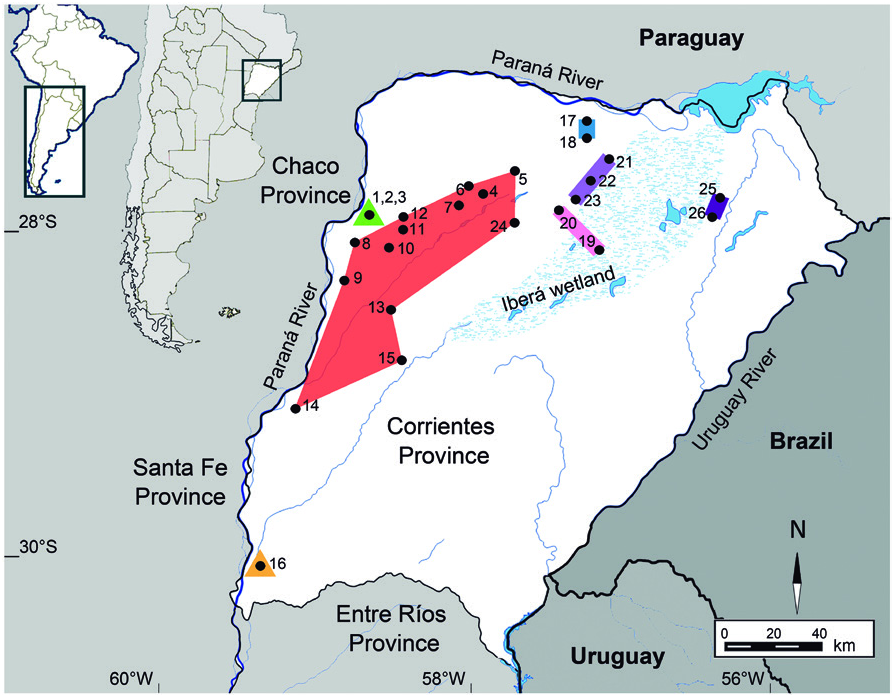
Ctenomys perrensi was initially described by Thomas (1896) based on a sample from Goya, its topotype. Afterwards, this population was ascribed to Ctenomys torquatus ( Cabrera 1961, Reig et al. 1965) and also to what would be later called Ctenomys pearsoni ( Langutth and Abella 1970) . During the decade of 1980 Contreras made a reassessment of the taxonomy and geographic range of Correntinean tuco-tucos, and extended the range for C. perrensi covering the central-west region of the province, from the margins of the Paraná River to the west margins of the Iberá wetland ( Figure 2 View Figure 2 ). This geographical range redefinition included all Correntinean populations except for the four nucleuses formerly ascribed to Ctenomys dorbignyi ( Contreras and Scolaro 1986, see above), the group of populations which were later alleged to Ctenomys roigi ( Contreras 1988) , and several unstudied populations between Goya and the former southern nucleus of C. dorbignyi ( Contreras et al. 1985) . However, the species range was further restricted by Ortells et al. (1990) to the topotype (Goya) and two populations that share the same karyomorph 2n = 50 FN = 84: Rincón de Ambrosio and Colonia 3 de Abril ( Table 1 View Table 1 ). Besides, a group of populations formerly attributed to this species was indicated to be probably related to this species based on morphological similarity and the sharing of a common FN = 84 ( Ortells et al. 1990). These highly polymorphic or polytypic populations, with 2n varying from 54 to 62, were: Saladas (and nearby locations), Mburucuyá (and nearby locations), San Roque, Yataytí Calle and M.F. Mantilla. Additionally, Ortells et al. (1990) acknowledged the membership of the population of Curuzú Laurel (2n = 42, FN = 76) to a different lineage since it had a clearly divergent karyotype. They suggested it should be related to C. torquatus , a species with similar 2n and FN. According to the IUCN Red List, C. perrensi is listed as Least Concern ( Bidau et al. 2008c), although its range of occupancy does not coincide with the one described by Contreras et al. (1985) or Ortells et al. (1990).
Under an integrative approach we delimit an extended lineage, the Ctenomys perrensi complex, encompassing all 2n <70 FN = 84 forms of the Corrientes group ( Table 1 View Table 1 ). These karyomorphs are the products of Robertsonian rearrangements ( Ortells et al. 1990, Lanzone et al. 2007, Caraballo et al. 2015), which not necessarily constitute reproductive barriers. In fact, Lanzone et al. (2007) showed that, in Correntinean tuco-tucos, heterozygotes bearing one or two centric fusion/fission rearrangements depicted normal pairing of homologous chromosomal arms. These results suggest that at least two Robertsonian changes would not act as a barrier to gene flow and can persist as intraspecific polymorphisms or polytypisms. However, heterozygotes for three or more centric fusions/fissions have not been reported in the Corrientes group ( Ortells et al. 1990, Lanzone et al. 2007, Caraballo et al. 2015), suggesting that highly Robertsonian-rearranged karyotypes are likely to produce hybrid sterility ( Baker and Bickham 1986). Karyomorphs with FN = 84 and 2n = 50 to 2n = 66 could form part of a chromosomal cline of neighboring interfertile or semi-interfertile forms, as occurs in the case of the Brazilian species Ctenomys minutus and Ctenomys lami ( Gava and Freitas 2003, Lopes et al. 2013).
To evaluate the Ctenomys perrensi complex from an integrative point of view, we should revise coincidences and discordances between SSR clustering and mtDNA phylogeny. The mtDNA phylogeny splits this complex into two clades: clade 2 including the populations of Ctenomys perrensi sensu Ortells et al. (1990) , Chavarría and Paraje Sarandicito ( Table 1 View Table 1 ) and clade 1, including the remaining populations as well as Ctenomys roigi which is, as discussed above, a different monophyletic lineage. The members of clades 1 and 2 of the C. perrensi complex resulted intermixed in SSR clusters ( Table 1 View Table 1 ). With the exception of cluster G, that groups Santa Rosa and San Miguel which are highly divergent populations in terms of their karyotypes and mtDNA identity and hence this grouping is likely to be artificial, all members of the C. perrensi complex fall into exclusive SSR clusters ( Table 1 View Table 1 ). Taken together, the mtDNA phylogeny and SSR clusters suggest these are recently diverged populations which share randomly assorted ancestral variants and/or are or have been genetically connected.
We consider Santa Rosa as a member of this complex, not only because it shares the same FN and is included in this group in the mtDNA phylogeny, but also because it falls into an exclusive cluster with all Ctenomys perrensi complex populations (San Roque, Chavarría and Goya) in previous SSR clusters ( Mirol et al. 2010). In turn, Paraje Sarandicito is not considered part of this lineage and will be discussed in the following item.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.