
Danakilia dinicolai, Stiassny, Melanie L. J., Marchi, Giuseppe De & Lamboj, Anton, 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.199529 |
|
DOI |
https://doi.org/10.5281/zenodo.5624407 |
|
persistent identifier |
https://treatment.plazi.org/id/03BD87B7-FFE7-222D-E1E6-F8B3BDA937E2 |
|
treatment provided by |
Plazi |
|
scientific name |
Danakilia dinicolai |
| status |
sp. nov. |
Danakilia dinicolai View in CoL , new species
Figs. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 ; Table 2 View TABLE 2
Holotype. MSNM 4888, male, 96.4 mm SL, Eritrea: Lake Abaeded. 14˚ 35' 17" N, 40˚ 05' 04" E., G. De Marchi, January 1998.
Paratypes. MSNM 4889, male, 72.9 mm SL, same data as holotype. MSNM 4890, female, 63.9 mm SL, same data as holotype. MSNM 4891, male, 56.7 mm SL, same data as holotype. AMNH 248881, 5 ex., 3 males, 2 females, 69.9–82.9 mm SL, Eritrea: Lake Abaeded, G. De Marchi, March 2001. AMNH 251633 4 ex. (1C&S), 1 male, 3 females, 63.6–76.5 mm SL, Eritrea: Lake Abaeded, G. De Marchi, March 2001. MSNG 55210, 1 female, 73.0 mm SL, Eritrea: Lake Abaeded. 14˚ 35' 17" N, 40˚ 05' 04", E., G. De Marchi, March 2001.
Diagnosis. Differs from its only congener, Danakilia franchettii , in the possession of longer pectoral fins (31.1–38.7% SL vs. 21.7–30.6% SL). Outer row teeth in both oral jaws composed of stout tricuspids, considerably more robust and fewer in number than those of D. franchettii . Lower pharyngeal bone relatively massive, longer than wide, length 30–32% HL (vs. small, only slightly longer than wide, length 25–27% HL in D. franchettii ), with 40–48 teeth in posterior row (vs. 26–36 in D. franchettii ).
Description. Based on holotype and 13 paratypes. Maximum size 96.4 mm SL. Counts and proportional measurements are given in Table 2 View TABLE 2 . Snout acute, mouth terminal, lips thickened but not fleshy. Head large, snout conical with straight profile, head smoothly convex behind nape to dorsal fin, except in largest males with a bulbous frontal gibbosity, particularly evident in life ( Figs. 2 View FIGURE 2 & 4 View FIGURE 4 B). Greatest body depth between insertion of dorsal and pelvic fins. Caudal peduncle always longer than deep. Caudal fin truncate. Dorsal XIII–XIV 9–11. Anal III 7 or 8. Pectoral fin long, reaching to anal fin origin or beyond. First pelvic ray longer in males than females but pelvic fin always reaching anus, and often beyond anal origin.
Jaws isognathous, both with 2–4 rows of tricuspid teeth. Outer row teeth in both jaws stout tricuspids with small, symmetrically rounded lateral cusps and larger medial cusp ( Fig. 3 View FIGURE 3 A), considerably more robust and fewer in number than in D. franchettii ( Fig. 3 View FIGURE 3 B). Lower pharyngeal bone relatively massive, longer than wide ( Fig. 3 View FIGURE 3 A), length 30–32% HL (versus small, only slightly longer than wide, length 25–27% HL in D. franchettii ( Fig. 3 View FIGURE 3 B). Pharyngeal teeth in anterior field of jaw, robust unicuspids, sparsely implanted, becoming increasingly elongate, recurved and spatulate posteriorly, densely implanted over posterior field of jaw, 40–48 teeth in posterior row ( Fig. 3 View FIGURE 3 A). Pharyngeal teeth in D. franchettii are similarly arrayed, but coarser and less densely implanted in posterior field of jaw, 26–36 teeth in posterior row ( Fig. 3 View FIGURE 3 B).
Eleven to 14 small, moderately tuberculate gill rakers along lower limb of first arch (including raker at angle), 2 or 3 pointed epibranchial rakers. Microbranchiospines absent.
Scales cycloid, 2 or 3 rows on cheek, 2–4 horizontal rows on opercle. Dark scaleless spot on outer edge of opercle. Chest and belly scales deeply embedded, markedly smaller than on flank, 4 or 5 scales between pectoral and pelvic fins. Upper lateral line separated from dorsal fin anteriorly by 3 or 4 scale rows, at 8th pored scale by 2 scale rows, at last pored scale by 1–1.5 scale rows. End of upper lateral line separated from lower lateral line by 1 or 2 scale rows. Basal 1/3 of caudal fin covered with scales; other fins unscaled.
holotype mean SD Range Standard length 96.4 77.9 63.6–96.4 Caudal peduncle length % as of depth 120.2 118.7 8.3 102.8–128.8
median
Gill rakers on lower limb of first arch 12 12 11–14 Total gill rakers on first arch 14 14 13–17
Miscellaneous osteology. Infraorbital series complete, with lachrymal (first infraorbital) and four tubular infraorbitals, lachrymal with five openings of laterosensory system. Total of 25 or 26 vertebrae, 11–13 precaudal and 12–14 caudal. Inferior vertebral apophysis (spondylophysial apophysis) supporting anterior end of gas bladder composed of elongate ventro-posteriorly directed process on 3rd vertebral centrum which abuts a stout ventro-posteriorly directed process on 4th centrum; similar in form and composition to that of Iranocichla ( Fig. 3 View FIGURE 3 D).
Coloration in life. Females and non-dominant males ( Fig. 4 View FIGURE 4 A). Opercule and body pale silvery-grey, snout and nape dark brownish-black. Dark scaleless spot on outer edge of opercle variously intense, but always present. Upper lip with bluish cast, lower lip grey. Throat and branchiostegal membranes pale silvergrey. Two dark longitudinal stripes sometimes visible on flanks, upper stripe at level of upper lateral line, passing to end of dorsal-fin base, lower stripe from edge of opercle to caudal base, not extending into caudal fin. Sometimes 5–7 dark vertical bars on body from dorsal-fin base to ventrum. Eye golden-yellow to brownish-black. Dominant males ( Fig. 4 View FIGURE 4 B). Dorsal, caudal and anal fins pale blue, with numerous paler blue dots. White margin of dorsal fin flushed pale red-orange distally. Thin white margin on upper caudal lobe. Pelvics blue, paler distally. Body scales with iridescent, pale blue posterior margins, centrally grey to greyishblack. Scales on chest and belly silvery grey, with darker grey margins. Prominent frontal gibbosity blackishblue to greyish-blue. Snout and lips pale blue.
Coloration in preservation ( Fig. 2 View FIGURE 2 ). Both sexes with head and body brown, darker dorsally, with dark lines or bars often visible as described in living specimens. Dorsal, anal and caudal fins heavily spotted in males, less so in females.
Feeding behaviour. No field observations made, but gut morphology and contents suggests a diet primarily consisting of algae grazed from the surface of plants and rocks. Guts are highly coiled, and extend to 7.6 times SL, most are filled with floculent algae with some sand grains and vegetable debris. However, it is likely that this species is an opportunistic feeder, as individuals were caught on lines baited with grasshoppers.
Breeding behaviour. Unknown, but large males guarding pits over sand were observed in the field.
Distribution and habitat ( Fig. 5 View FIGURE 5 ). Lake Abaeded occupies a small oval depression (maximum width of 450 m, maximum depth 10 m) in a volcanic crater at the northwestern border of the Danakil Depression in Eritrea, 132 km southeast of the coastal town of Massawa. Lake surface is about 30 m below sea level. Sand from dunes along the eastern and southern rim is frequently blown into the lake. The lake is fed by many hot springs at surface level and underwater, and as there is no surface outlet evaporation in this hyper arid region (rainfall of ~ 5 m /year) probably fully compensates for the input from springs to maintain a stable lake level year round. In April 1998 water temperature was 29°C but reached 45°C close to the hot springs. Total salinity was 14.5 gr/l. The most abundant ions were Na+ (111 EPM), Ca++ (99 EPM), Mg++ (18 EPM), Cl - (217 EPM) and SO4 - (41 EPM). Ph was 7.46 and EC was 23100. High concentrations of Na+, Ca++, Cl - and SO4 - are certainly the result of circulation of water heated at depth through layers of gypsum and halite that cover the bottom of the northern part of the Danakil Depression ( Gasse & Street 1978). The lakeshore is almost completely encircled by reeds ( Phragmites australis ) and stands of tamarisk ( Tamarix aphylla ), but is free of vegetation in the proximity of hot springs. A single non-piscine vertebrate inhabiting the lake is a frog, probably Ptychadena anchietae . Fishes are abundant in the lake and concentrate among the reeds, but are found also on sandy substrate near the shore, where temperature is not too high. The bottom of the sandy areas is covered with crater-like nests dug and defended by large colourful males. The species has been found so far only in Lake Abaeded, but no research has yet been conducted in the nearby Lake Badda, a variable surface reed-fringed lake only 8 km north of Lake Abaeded ( Fig. 5 View FIGURE 5 ).
Etymology. Named in memory of Ernesto Di Nicola, a member of the 2001 expedition to Lake Abaeded who died in a car accident while returning from the lake.
Discussion. The discovery of a second species of Danakilia in a small lake currently isolated by some 160 km of hyper-arid desert from its congener, Danakilia franchettii , raises the question of the age and origin of these two isolated species. The geological history of the region indicates that the Danakil Depression formed as a result of tectonic movements related to the Afro-Arabian Rift ( Beyene & Abdelsalam 2005). It was an intermittent marine basin (at least in the northern region) during the late Pleistocene, and with the formation of volcanic structures at the northern end it is thought to have separated from the Red Sea sometime around 32,000–25,000 years ago ( Bonatti 1971, Barberi et al. 1972). Separation of the two species of Danakilia might therefore date back to the Pleistocene to the time of the formation of the Alayta e Tat'Ale shield volcanoes that surround Lake Afdera ( Barberi et al. 1972). Alternatively, their separation may be considerably younger having occurred after the drying of extensive saline lakes that occupied the bottom of the Danakil Depression during a major arid phase that started about 9,000–5,000 years ago ( Gasse 1974, Gasse & Street 1978). Unfortunately, despite considerable effort all attempts to extract DNA from either species have so far failed, and molecular divergence time estimates cannot be made until additional material becomes available.

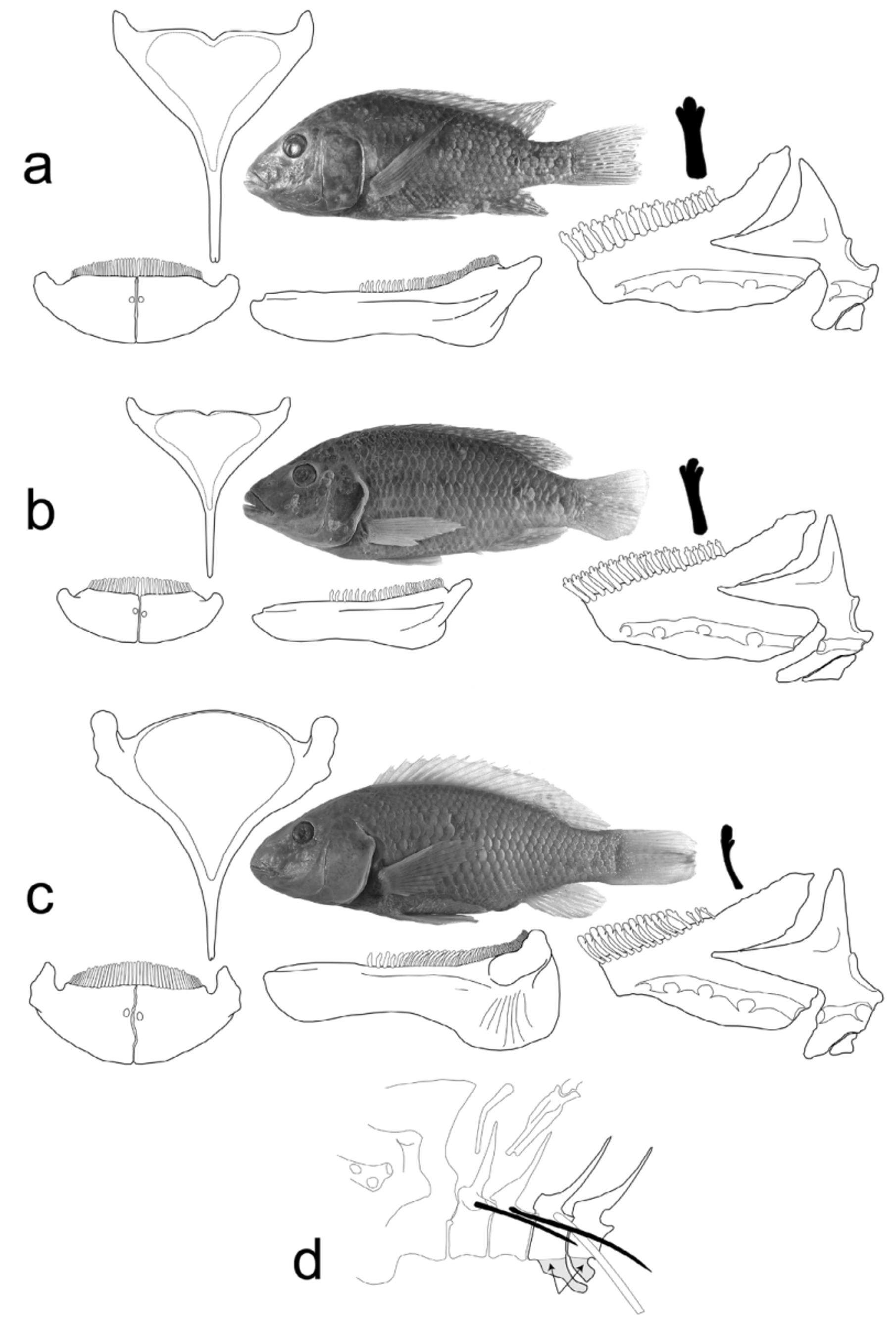

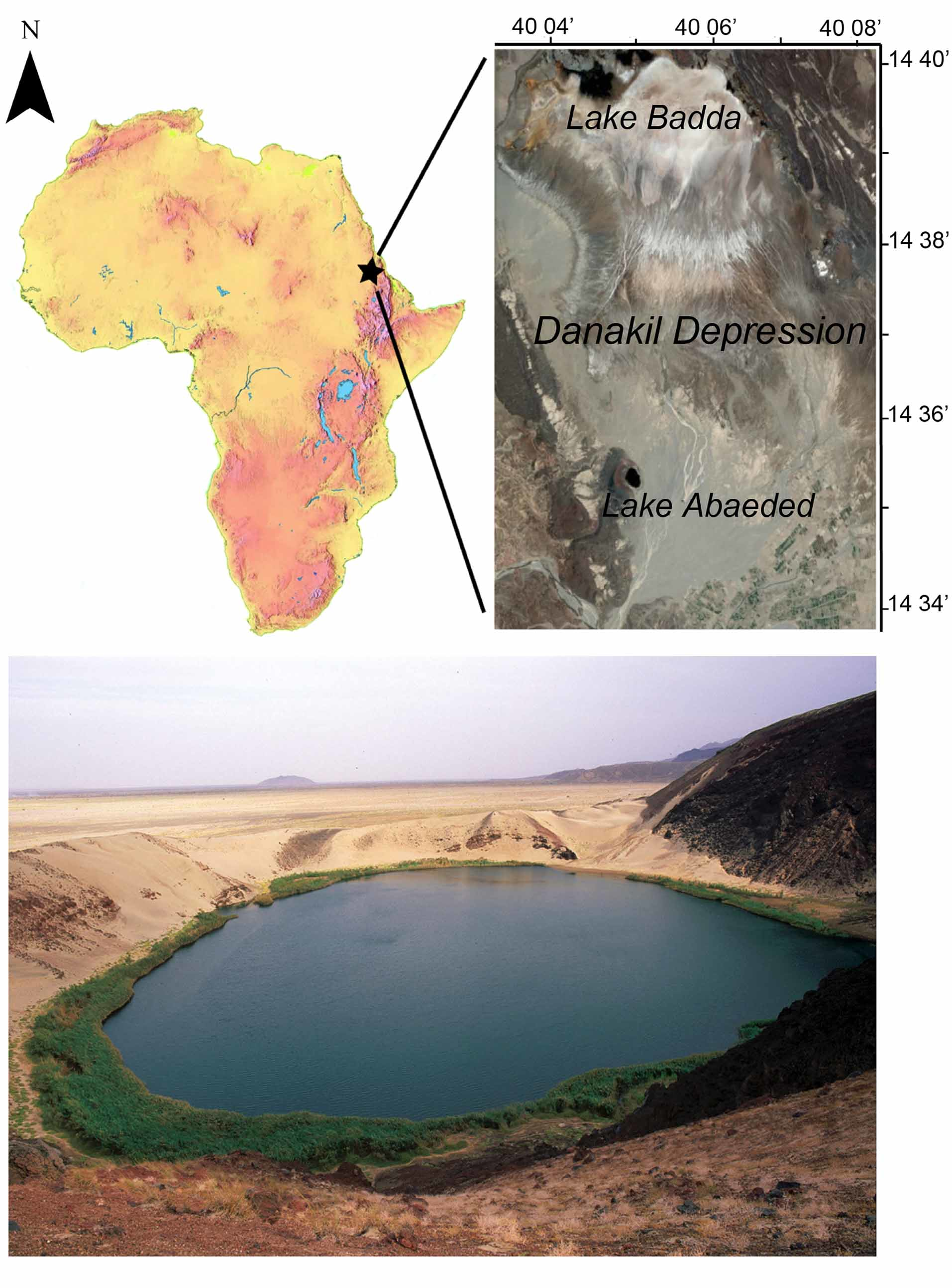
TABLE 2. Morphometric and meristic data for the holotype and 13 paratypes of Danakilia dinicolai.
| % SL | |||
|---|---|---|---|
| Body depth | 41.0 | 39.4 | 1.2 37.2–41.3 |
| Head length | 39.2 | 38.9 | 1.2 37.0–40.9 |
| Caudal peduncle length | 14.2 | 14.2 | 0.9 12.5–15.2 |
| Caudal peduncle depth | 11.8 | 11.9 | 0.5 11.4–13.4 |
| Dorsal-fin base | 49.2 | 48.7 | 1.3 46.8–51.0 |
| Anal-fin base | 13.7 | 13.5 | 1.0 12.0–14.9 |
| Predorsal distance | 39.2 | 40.0 | 1.0 38.7–41.7 |
| Preanal distance | 75.7 | 73.3 | 2.6 69.5–76.3 |
| Prepectoral distance | 44.5 | 42.1 | 1.0 40.8–44.5 |
| Prepelvic distance | 47.4 | 45.3 | 1.3 43.5–47.4 |
| Longest dorsal-fin ray | 28.3 | 17.1 | 3.9 13.2–28.3 |
| Longest anal-fin ray | 25.4 | 16.1 | 3.1 13.8–25.4 |
| Longest pectoral-fin ray | 31.1 | 33.6 | 2.5 31.1–38.7 |
| Longest pelvic-fin ray | 32.6 | 24.0 | 2.9 21.2–32.6 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |