
Amphisbaena mebengokre, Ribeiro & Sá & Santos-Jr & Graboski & Zaher & Guedes & Andrade & Vaz-Silva, 2019
|
publication ID |
https://doi.org/10.11646/zootaxa.4550.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:A493A2CB-5084-42A2-B64F-5B4526FDC273 |
|
DOI |
https://doi.org/10.5281/zenodo.5935213 |
|
persistent identifier |
https://treatment.plazi.org/id/03BE1739-FFB2-FFB8-63BC-B093FBC8F967 |
|
treatment provided by |
Plazi |
|
scientific name |
Amphisbaena mebengokre |
| status |
sp. nov. |
Amphisbaena mebengokre sp. nov.
( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 , 3 View FIGURE 3 ; Tables 1, 2)
Holotype. An adult male ( MNRJ 25189 View Materials ) collected on the banks of the Caiapó River ( 16 o 27’25’’S, 51 o 22’41’’W; 464 m above of sea level; datum = WGS84 ) near to the Hydroelectric Power Plant Santo Antônio do Caiapó , Municipality of Arenópolis, State of Goiás, Brazil, on 22 November 2012 by Jafi M. Carmo and Wilian Vaz-Silva. GoogleMaps
Paratypes. Adult males: MNRJ 25191 View Materials , MNRJ 25194 View Materials , MNRJ 25196 View Materials , MNRJ 25198 View Materials , MPEG 32206 View Materials , ZUFG 1194 , ZUFG 1196 , ZUFG 1201 , ZUFG 1204 , ZUFG 1205 , ZUFG 1207 . Adult females: MNRJ 25190 View Materials , MNRJ 25192 View Materials , MNRJ 25193 View Materials , MNRJ 25195 View Materials , MNRJ 25197 View Materials , MPEG 32205 View Materials , ZUFG 1193 , ZUFG 1195 , ZUFG 1197 , ZUFG 1200 , ZUFG 1202 , and ZUFG 1203 . Undetermined: ZUFG 1206 . All paratypes are from the type locality, Arenópolis municipality, state of Goiás, Brazil, collected by Wilian Vaz-Silva, Jafi M. Carmo, Paulo Roberto Gomes, and Edmar P. Victor between 22 and 28 November 2012 .
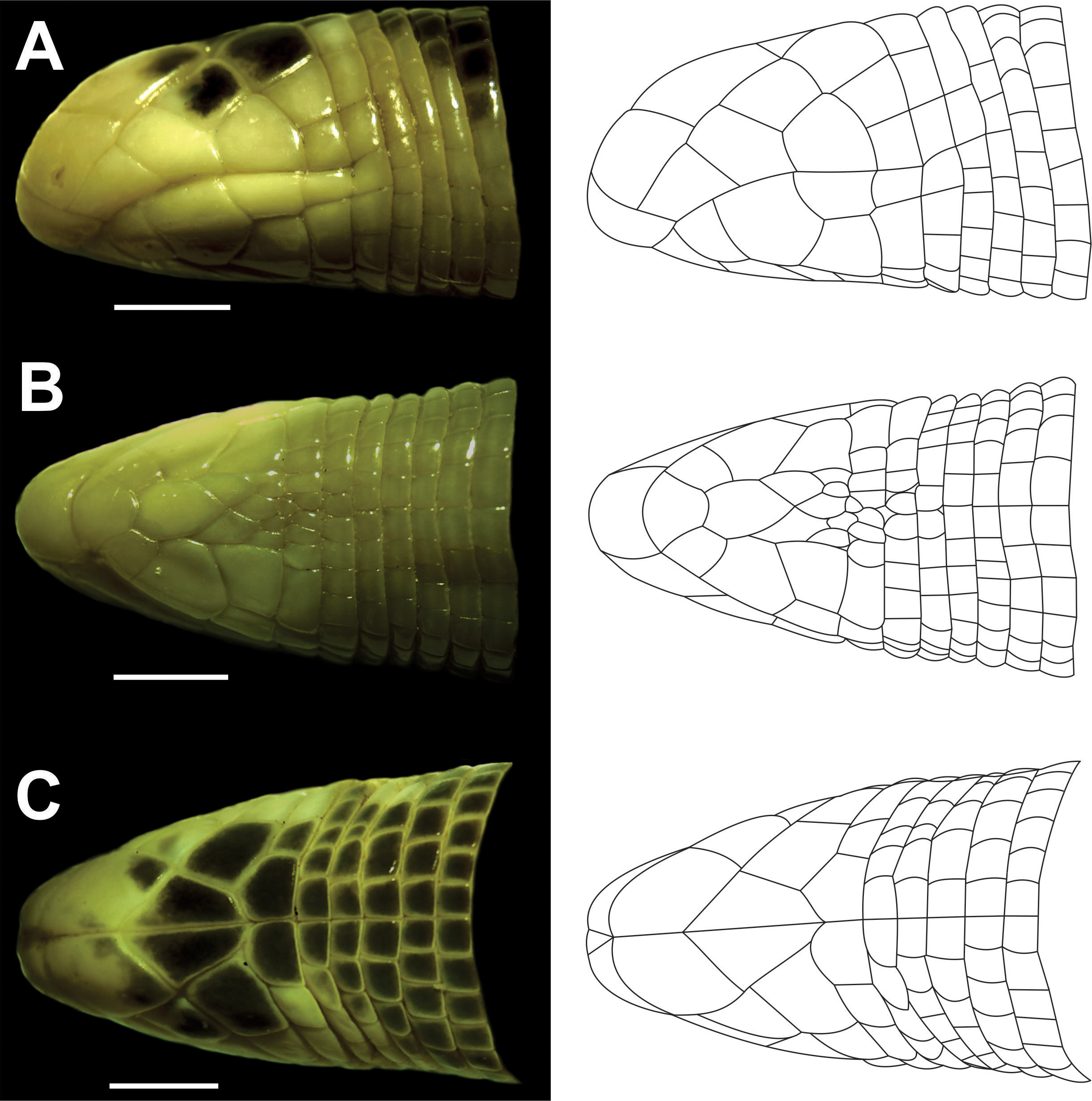
Diagnosis. The new species is a member of the Amphisbaena silvestrii group ( sensu Vanzolini 1997) and is distinguished from all congeners by the combination of the following characters: (1) snout rounded in dorsal view and slightly convex in profile view; (2) two precloacal pores; (3) 161–176 dorsal half-annuli; (4) 12–15 tail annuli; (5) 3/3 supralabials; and (6) postmalar row absent.
Comparisons with other species. Amphisbaena mebengokre sp. nov. can be distinguished the other South American amphisbaenians species by a combination of characters. Differs from Amphisbaena acrobeles ( Ribeiro, Castro-Mello & Nogueira, 2009) ; Amphisbaena anomala ( Barbour, 1914) ; Amphisbaena bilabialata ( Stimson, 1972) ; Amphisbaena kingii ( Bell, 1833) ; Leposternon bagual Ribeiro, Santos-Jr & Zaher, 2015 ; Leposternon cerradensis Ribeiro, Vaz-Silva & Santos-Jr, 2008 ; Leposternon infraorbitale ( Berthold, 1859) ; Leposternon kisteumacheri Porto, Soares & Caramaschi, 2000 ; Leposternon maximus Ribeiro, Nogueira, Cintra, Silva Junior & Zaher, 2011 ; Leposternon microcephalum Wagler, 1824 ; Leposternon mineiro Ribeiro, Silveira & Santos-Jr, 2018 ; Leposternon octostegum ( Duméril & Duméril, 1851) ; Leposternon polystegum ( Duméril & Duméril, 1851) ; Leposternon scutigerum ( Hemprich, 1820) ; Leposternon wuchereri ( Peters, 1879) ; Mesobaena huebneri Mertens, 1925 ; and Mesobaena rhachicephala Hoogmoed, Pinto, Rocha & Pereira, 2009 mainly by having snout rounded in dorsal view and slightly convex in profile view (vs. snout shovel-like or compressed forming a sharp and prominent keel) (details of cited species in original descriptions and studies of Gans 1971a, 1971b; Mott et al. 2008; Hoogmoed et al. 2009; Ribeiro et al. 2009; Ribeiro 2010; and Ribeiro et al. 2018). It differs from all others Amphisbaena species with rounded snout, except of A. absaberi , A. anaemariae , A. brevis , A. caiari , A. carli , A. crisae , A. cuiabana , A. darwini , A. dubia , A. filiformis , A. hiata , A. kiriri , A. leeseri , A. lumbricalis , A. miringoera , A. mitchelli , A. neglecta , A. persephone , A. roberti , and A. silvestrii by having two precloacals pores. The new species differs from all Amphisbaena species with two precloacals pores, except A. anaemariae , A. crisae , A. darwini , A. kiriri and A. silvestrii by having 161–176 dorsal half-annuli (vs. less than 140 dorsal half-annuli in A. brevis , less than 154 in A. neglecta , and more than 188 dorsal half-annuli in the other species). It differs from Amphisbaena anaemariae , A. crisae , A. darwini , and A. silvestrii by having 12–15 tail annuli (vs. more than 18 tail annuli in the other species); and A. kiriri by having 3/3 supralabials and postmalar row absent (vs. 4/4 supralabials and postmalar row present).
Description of the holotype. A male adult; snout-vent length 159 mm; tail length 12 mm; midbody diameter 6.7 mm (4.2% of snout-vent length); head length 5.7 mm (ca. 3.6% of snout-vent length) distinct from the neck (slightly constrict); snout rounded in dorsal view and slightly convex in profile view; rostrum projecting forward beyond the jaw.
Rostral slightly visible in dorsal view ( Fig. 1C View FIGURE 1 ), subtriangular in ventral view ( Fig. 1B View FIGURE 1 ), lateral portion straight, in contact with nasals laterally and first supralabial latero-posteriorly. Nasals with irregular shape, paired, with a small middorsal suture (suture length ca. 14.1% of head length), posterior portion straight, in contact with first supralabial laterally, second supralabial latero-posteriorly and prefrontals posteriorly. Nostril near the anteroinferior angle of the nasal shield ( Fig. 1A View FIGURE 1 ). Prefrontals paired, relatively large (ca. 30% of head length), with a long middorsal suture (ca. 30% of head length), longer than the nasal middorsal suture, shorter than the frontal middorsal suture (ca. 42% of head length), anterior portion convex, latero-posterior portion projected, in contact with second supralabial and ocular laterally, frontals posteriorly and in point contact with post-ocular posteriorly. Frontals irregularly polygonal, paired, with a long middorsal suture (ca. 30% of head length), elongated, 1.9 times longer than wide, separated from ocular by the point contact of the prefrontal and postocular, in contact with postocular laterally and parietal posteriorly. Parietals irregularly polygonal, paired, with a short middorsal suture (ca. 12% of head length), almost as long as wide, with their posterior border straight and parallel to the second body annulus, in contact with postocular laterally, and second body annulus posteriorly. The anterior edge of the parietals is at the level of the angulus oris. Body annulus behind parietals, shows 25 regular middorsal scales ( Fig. 1C View FIGURE 1 ). Ocular diamond-shaped, relatively long (length representing ca. 30.2% of head length), in broad contact with second and third supralabials laterally and postocular posteriorly. Eyes visible, located in near anterior angle. Postocular pentagonal, relatively long (length. 27.7% of head length), almost as high ( 1.8 mm) as long ( 1.6 mm), in lower contact with temporal, with their posterior border straight and parallel to the second body annulus posteriorly ( Fig. 1A View FIGURE 1 ).
Three supralabials, irregularly polygonal, increasing in size posteriorly; first rhomboid, smallest, 1.1 times longer than high, in contact with second supralabial posteriorly; second supralabial relatively high and narrow, forming an irregular trapezium, ca. 0.8 times higher than wide, in contact with frontal third supralabial posteriorly; third supralabial pentagonal, almost as high ( 1.3 mm) as wide ( 1.2 mm), in contact with temporal and postsupralabial posteriorly. Posteriorly to the supralabials there is a row of scales that includes the postocular, temporal and postsupralabial. Temporal smaller than postocular, limited to the second body annuli posteriorly, in point contact with ocular, in upper contact with the postocular and with the upper part of the third supralabial, in lower contact with postsupralabial. One quadrangular postsupralabial, relatively small (ca. 12.3% of head length), limited to the second body annuli posteriorly.
Three infralabials, first irregular, in point contact with postmental, in broad contact with mental and second infralabial. Second infralabial irregularly polygonal, largest (length representing ca. 27.5% of head length), longer than wide, in contact with postmental and genials laterally and malar and third infralabial posteriorly. Third infralabial smallest, nearly rectangular, longer than wide, in contact with malar laterally and first body annulus posteriorly. On the side of the third infralabial, is a slightly enlarged scale of first body annulus (behind angulus oris) that appears to continue the infralabial series. Mental single, anteriorly abruptly widening, only slightly larger than postmental in the posterior border, lateral borders straight, in broad contact with postmental posteriorly. Postmental single, relatively long (26.5%), pentagonal, in broad contact with first row of postgenials posteriorly. Two rows of postgenials; first with two distinctly larger scales and one smaller, in contact with malars laterally and with second postgenial row posteriorly. Second postgenial row with five irregularly polygonal equally sized shields, in contact with malars laterally and first body annuli posteriorly. Two malars, one on each side, irregularly polygonal, in contact with first body annulus posteriorly. Postmalar row absent.
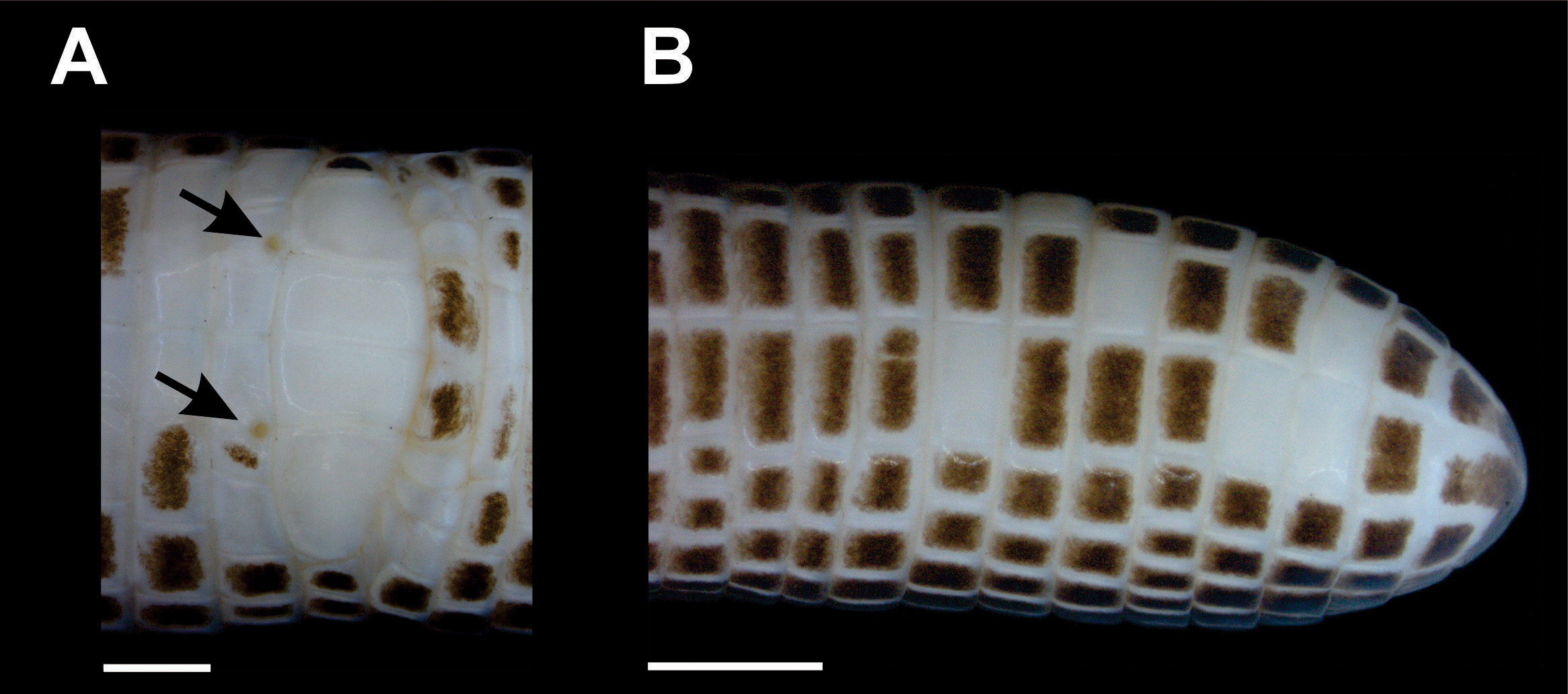
Body annuli distinct. The first body annulus is well-defined only ventrally. Dorsally, the first body annulus includes the postsupralabial, temporal and the very large postocular, dorsally limited by the frontal and parietal. Body annuli 170 with ventral part missing. Two lateral annuli in the cloacal region. Dorsal and ventral sulci absent; 14 dorsal and 12 ventral segments per midbody annulus; ventral segments with two central scales larger than adjacent scales and, only these, slightly larger than dorsal scales. Lateral sulci well marked and apparent from the 31 st body annulus almost to level of cloaca. Two precloacal pores in two adjacent central scales of the precloacal shield ( Fig. 2A View FIGURE 2 ). Cloacal segments regular, four larger and one small and 10 postcloacal segments. Fourteen caudal annuli; fifth tail annulus is slightly narrowed, it may correspond to an autotomic site ( Fig. 2B View FIGURE 2 ). Tip of tail rounded ( Fig. 2B View FIGURE 2 ).
Pattern color on preservative of the holotype. Anterior portion of head whitish, posterior portion of prefrontals, frontals, ocular, postocular and parietals dark brown ( Fig. 4 View FIGURE 4 ). Trunk and tail are uniform dark brown. Venter of head, mid of belly and cloaca whitish, lateral of belly and ventral portion of tail present a checkerboard pattern with dark brown segments.
Variation. Morphometric and meristic variations of the type series are given in Table 1. A few variations in the position of the cephalic shields were observed, when compared to the holotype. The paratypes ZUFG 1197 and ZUFG 1207 present parietal in a rectangular shape. ZUFG 1198 presents an azygous shield between the right parietal and first annulus of the body. ZUFG 1203 have a rectangular parietal only on the right side of the body. MNRJ 25191 presents an azygous shield between the parietal shield and the frontal on the right side of the body. MNRJ 25196 has irregular segments in part of the dorsal portion of the first annuli of the body.
Etymology. Amphisbaena mebengokre sp. nov. was found on the banks of the Caiapó River. The river's name gives homage to the indigenous people who inhabited the region and were expelled by the colonizers. The term "Kayapó" was first used in the early nineteenth century. The term was created by neighboring indigenous groups and means "those who look like monkeys" (a reference to a ritual using monkey masks). The Kayapó people prefer to call themselves "Mebêngôkre", which means "men of the springs." The specific epithet mebengokre gives homage to the Mebêngôkre ethnicity.
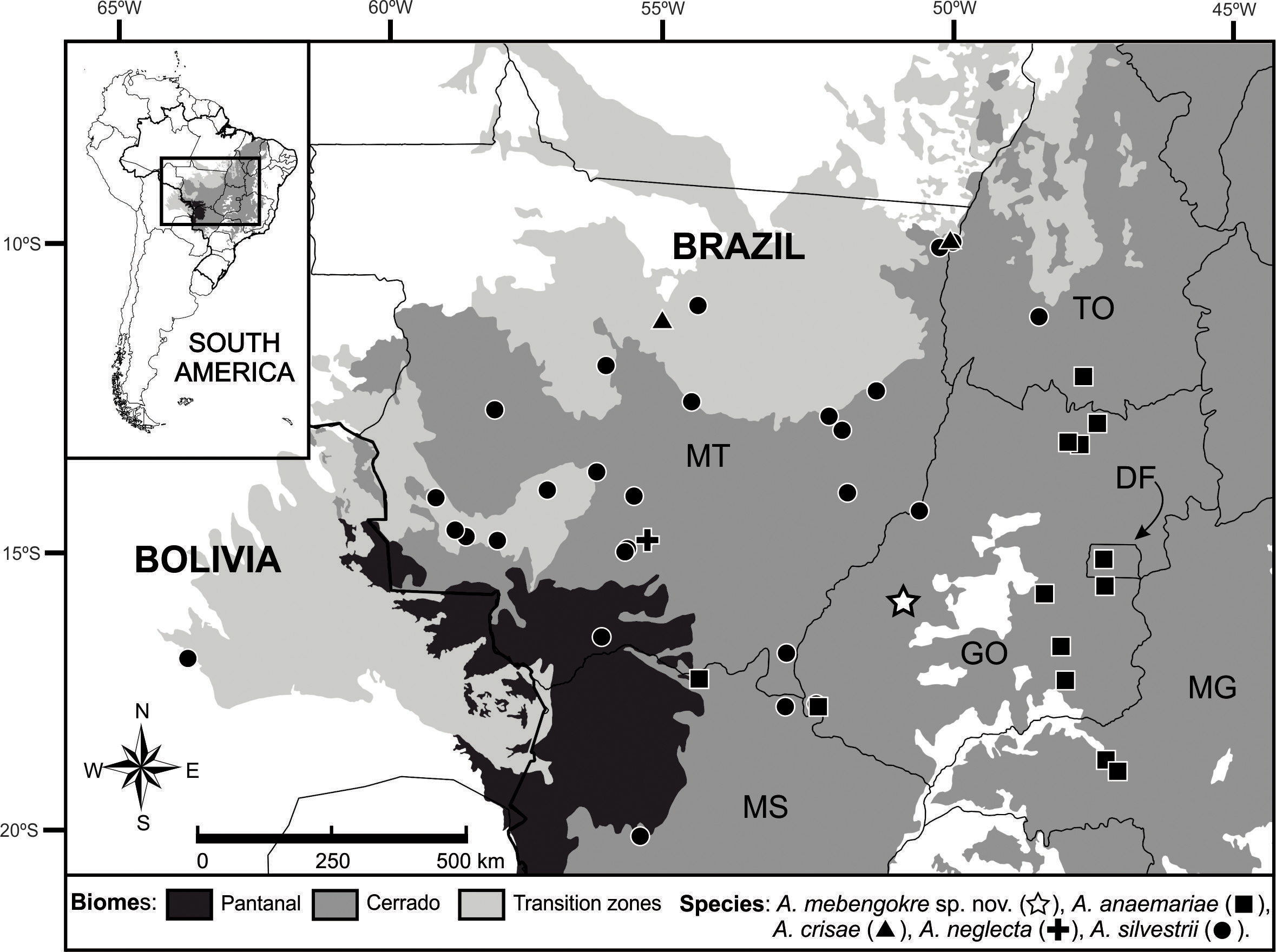
Distribution and habitat. Amphisbaena mebengokre sp. nov. is know only to the type locality in the municipality of Arenópolis, core region of the Cerrado biome in western state of Goiás ( Fig. 4 View FIGURE 4 ) The region is drained by the Caiapó River, a tributary of the right bank of the Araguaia River. The regional landscape is characterized by phytophysiognomies that include forest and savanna categories (Typical Cerrado, Deciduous Dry Forest, Riparian Forest and Gallery Forest). The type series was collected during the reservoir formation of the Hydroelectric Power Plant Santo Antonio do Caiapó, in Riparian Forest remnants. These remnants are limited to narrow bands of primary vegetation along the Caiapó River, and in some cases disappear and give way to savanna formations or have been replaced by pastures.
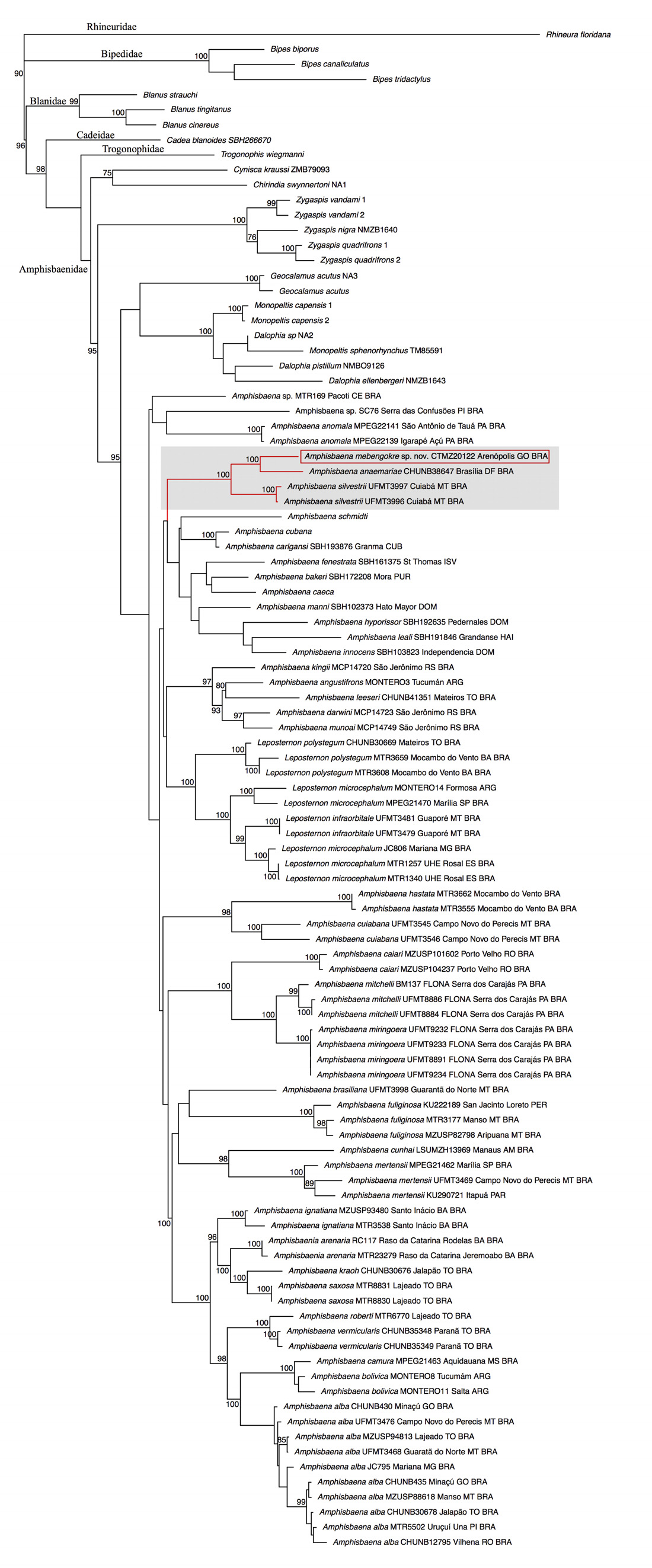
Phylogenetic relationships. Our concatenated alignment for five genes resulted in a matrix with a total of 3.966 base pairs. The proportion of gaps and completely undetermined characters in this alignment was 38.6%. The Partition Finder identified 11 partitions with GTR+G model for maximum Likelihood. The maximum likelihood analyses found a single tree with log likelihood of -42635.245032. The general topology resulted of the maximum likelihood analyses ( Fig. 5 View FIGURE 5 ) was similar to previous works, as Vidal et al. (2008) and Mott & Vieites (2009), for the mains groups of Amphisbaenia . Amphisbaena mebengokre sp. nov. was recovered as a sister group of A. anaemariae with 100% of bootstrap support. The clade formed by A. mebengokre sp. nov. and A. anaemariae was recovered as sister group of A. silvestrii with high value of bootstrap support (100%). The genetic distance ( pdistance) between A. mebengokre sp. nov. and A. anaemariae is 16.5%, and between A. mebengokre sp. nov. and A. silvestrii is 18.9% for ND2 gene; and the genetic distance ( p -distance) between A. mebengokre sp. nov. and A. anaemariae is 6.9%, and between A. mebengokre and A. silvestrii is 7.8% for 16S gene.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |