
Leucostethus bilsa, Vigle & Coloma & Santos & Hernandez-Nieto & Ortega-Andrade & Paluh & Read, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4896.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:383BE990-AF74-461B-8449-F41CCCDC94CD |
|
DOI |
https://doi.org/10.5281/zenodo.4389609 |
|
persistent identifier |
https://treatment.plazi.org/id/03BE87C4-FFAC-FFA4-62F2-0B8BED84FA02 |
|
treatment provided by |
Plazi |
|
scientific name |
Leucostethus bilsa |
| status |
sp. nov. |
Leucostethus bilsa sp. nov
Colostethus View in CoL sp. Ortega-Andrade,Bermingham, Aulestia, and Paucar (2010b), Check List 6: 119–154.
Proposed standard English name. Bilsa white-chested frog
Proposed standard Spanish name. Rana pecho blanco de Bilsa
( Figs. 2–10 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 , 12D View FIGURE 12 )
Holotype. ( Figs. 2 View FIGURE 2 , 5F, G View FIGURE 5 , 8 View FIGURE 8 , 12D View FIGURE 12 ). CJ 8311 , an adult male collected in a primary forest at the Brown Trail (0.358389 N, 79.708364 W; 478 m) in Reserva Biológica Bilsa ( RBB), Quinindé, Provincia de Esmeraldas, Ecuador, 21 March 2018, by Sebastián Hernández-Nieto. GoogleMaps
Paratypes. ( Figs. 3B View FIGURE 3 , 4A, C, D, F View FIGURE 4 , 5 View FIGURE 5 A–E, 6A–D, 7A–C, 9) 22 preserved frogs (5 adult females, 4 adult males, 13 non-adults): QCAZ 14699 View Materials (gravid adult female) , QCAZ 14700 View Materials (adult male) both collected between the small streams known locally as the Río Aguacatal and the Río Rompe Frente in the RBB, ~ 2 km SW of the Bilsa biological station ( BBS) central facilities complex (0.355833 N, 79.703333 W, ~ 450 m elevation, Provincia de Esmeraldas, Ecuador, on the afternoon of 6 July 2000, by Russell G. Burge and William H. Schaedla GoogleMaps ; MCZ A-140205 (adult female), KU 327677 (sub-adult female) collected by day from areas ~ 3–4 km SW of the RBB on 15 July 2000 , by a group of Bilsa park guards (collected for Gregory O. Vigle); CJ 7765 (female, SVL = 22.5 mm) , CJ 7775 (female, SVL = 25.6 mm, C&S specimen), collected as juveniles and raised to adult stage in the lab, from Río Rompe Frente in the RBB on 20, 21 January 2018 by Sebastián Hernández-Nieto; CJ 8322 (female, SVL = 28.2 mm) collected from the RBB on 13 May 2018 ; CJ 8275 and CJ 8299 (post-metamorphs) collected from the Duchas stream (0.345083 N, 79.712347 W; 423 m) of the GoogleMaps RBB on 22 February 2018 , and 20 March 2018 respectively by Sebastián Hernández- Nieto ; CJ 8416 (post-metamorph) collected from the White trail (0.348086 N, 79.716569 W; 426 m) of the GoogleMaps RBB on 11 June 2018 by Sebastián Hernández-Nieto; CJ 8198 , 8201 , 8202 , 8204 , 8274 , 8334 , and 8413 (7 juveniles) collected from the RBB on 4, 5 December 2017, 22 February 2018, 15 March 2018 and 11 June 2018 by Sebastián Hernández-Nieto; KU 327679 (adult male carrying larvae) collected by day ~ 2.3 km SW of the BBS on 16 August 2002 , by Scott D. Cashins ; INABIO 3632 (adult male carrying larvae) collected ~ 1 km SW of the BBS on 11 December 2006 ; INABIO 3649 (sub-adult female) collected ~ 2 km SW of the BBS on 13 December 2006 ; INABIO 3785 (sub-adult female), and INABIO 3786 collected ~ 2 km SW of the BBS on 26 February 2007 , by H. Mauricio Ortega-Andrade .
Etymology. The specific epithet is used as a noun in apposition and refers to the type locality, the Reserva Biológica Bilsa; Bilsa is the name of a nearby river. RBB protects the headwaters of several rivers and belongs to the Chocó Darien-Tumbesino region, one of 25 global “hot spots” known for their high biodiversity and endemism of flora and fauna. This name is also intended to honor the conservation efforts of Fundación Jatun Sacha, which owns and administers the RBB and several other reserves in Ecuador.
Definition. Leucostethus bilsa is characterized by the following combination of characters: (1) large sized, snout-vent length (SVL) in females 27.4–28.2 mm (n = 3, wild caught), 22. 5–28.0 (n = 3, lab-raised); SVL in males 21.7–24.2 mm (mean = 22.7; n = 4); (2) skin of dorsum posteriorly tubercular, smooth anteriorly, but with scattered low tubercles on the lateral sides of the dorsum anteriorly; (3) Finger I slightly longer than Finger II when adpressed; (4) length of Finger IV reaching distal half of distal subarticular tubercle of Finger III; (5) Finger III not swollen in adult males; (6) lateral fringes absent on fingers; (7) discs on fingers weakly to moderately expanded; (8) webbing absent between fingers; (9) outer tarsal fold absent, inner tarsal fold well-developed; (10) lateral fringe absent on Toe IV, but keels are present in toes II–IV; (11) discs on toes weakly to moderately expanded; (12) basal webbing between toes III–IV–IV, only trace webbing between toes II–III; (13) ventrolateral stripe absent, but junction of dark lateral with pale ventral surfaces abrupt, forming a sharply distinct linear border; (14) cream oblique lateral stripe present, usually incomplete; (15) in life, mustard-yellow paracloacal marks present; (16) in life, bright mustard-yellow flash marks in axillae, groin, and antero-ventral surfaces of hindlimbs; (17) in life, ventral surfaces of body white with yellow in posterior portion of belly, with gray mottling on chin; throat, and chest in adult males; (18) testis color polymorphic, white or brown-gray; (19) thin distinct dark stripe or flecks present along lower lip; (20) advertisement call composed of a series of peep notes emitted at a mean rate of 81 notes/minute, with dominant frequency of 3310–3722 Hz.
Comparisons with other species. Leucostethus bilsa differs from all Leucostethus (except L. argyrogaster ) by lacking the Finger III swollen in adult males. Leucostethus bilsa is compared with all other Leucostethus described ( L. argyrogaster , L. brachistriatus ( Rivero & Serna 1986) , L. fraterdanieli ( Silverstone 1971) , L. fugax , L. jota Marin et al. 2018 , and L. ramirezi ( Rivero & Serna 2000) , and similar species of Dendrobatidae occurring in the lowlands of northwestern Ecuador ( Hyloxalus toachi ( Coloma 1995) , H. awa ( Coloma 1995) , H. infraguttatus ( Boulenger 1898)) . Leucostethus bilsa is distinguished from L. argyrogaster by having bright mustard-yellow flash marks (absent in L. argyrogaster ). Leucostethus bilsa differs from L. fraterdanieli (characters in parenthesis) in that the flash marks have diffuse borders (more defined), and form crescent-shaped arcs around arm insertions, extending to ventral surfaces (do not consistently form crescent-shaped arcs). Also, L. bilsa differs in having a complete to incomplete oblique lateral stripe (prominent and fully complete oblique lateral stripe), a dark line below oblique lateral stripe is absent (the oblique lateral stripe is bordered below by a distinct dark line). Ventrolateral patterns also differ; while both species lack a pale ventrolateral stripe, the border between the dark flanks and pale ventral surfaces is abrupt and linear in L. bilsa , forming a sharply defined straight or slightly sinuous line (fragmented). Dorsally (in life) both species are cryptically colored, with L. bilsa varying from brown to charcoal (dorsum may also have green, orange or golden tinges). Leucostethus bilsa further differs from L. brachistriatus , L. fugax , L. jota , and L. ramirezi by having an oblique lateral stripe that varies from complete to anteriorly faded or broken along its posterior length (pale oblique lateral stripe complete in L. brachistriatus , L. fugax , L. jota , and L. ramirezi ). Additionally, it differs from L. jota by having mustard-yellow paracloacal marks (paracloacal marks absent in L. jota ) and by lacking a ventrolateral stripe (ventrolateral stripe present in L. jota ). Leucostethus bilsa is larger than L. ramirezi (maximum female SVL = 28.0 mm vs 22.8 mm in L. ramirezi ).
Leucostethus bilsa and Oophaga sylvatica ( Funkhouser 1956) are the largest species of dendrobatid frogs yet reported from western Ecuador; only in Hyloxalus toachi does the size of adults of either sex approach or exceed the sizes of adults in the type-series of L. bilsa (maximum sizes of adult males and females respectively for H. toachi are 23.1 and 28.2 mm SVL). Leucostethus bilsa may readily be distinguished from H. toachi , H. awa , and H. infraguttatus by having bright mustard-yellow axillary and groin flash marks, and presence of mustard-yellow paracloacal marks (absent in H. toachi , H. awa , and H. infraguttatus ).
Acoustic comparisons. The call of Leucostethus bilsa (see description later under Vocalization) is compared to the calls of L. fraterdanieli and L. jota described by Marin et al. (2018). The calls of L. brachistriatus and L. ramirezi are unknown. The calls of all three species consist of a long string of roughly equally spaced notes, but the calls mostly differ in the call repetition rate of 98 notes per minute for L. fraterdanieli , 87 for L. bilsa , and 22 for L. jota . Leucostethus bilsa and L. jota produce much longer notes, note duration being 139 ms in L. bilsa , 140 ms in L. jota vs. 90 ms in L. fraterdanieli . The dominant frequency in L. fraterdanieli is 3.1–3.5 Khz, whereas the L. bilsa (3.6–3.7 KHz) and of L. jota (3.7–4.9 Khz) are higher.
Description of holotype. CJ 8311, an adult male, 24.17 mm SVL; body robust; head longer than wide, head width 32% of SVL, head length 35.91% of SVL; snout short, truncate and acuminate at the mid-line in dorsal view, bluntly rounded and protruding slightly beyond lower jaw in profile. Loreal region slightly concave; canthus rostralis smoothly rounded, moderately well-defined; nostrils protuberant and directed laterally. Supra-tympanic fold evident but not pronounced, obscuring postero-dorsal portion of tympanic annulus; tympanum prominent, 49.26% of eye length, tympanic annulus prominent, most pronounced on antero-ventral 1/3; tympanum 0.56 mm from eye at closest point.
Forelimbs moderately slender, skin smooth, ulnar fold absent. Finger I slightly longer than finger II when adpressed; relative length of fingers III>I>II>IV. Webbing, lateral fringes, and keels absent on hands and fingers; Finger III not swollen. Palmar tubercle large, irregularly rounded; thenar tubercle elliptical, about 1/2 the size of palmar tubercle; supernumerary tubercles absent; subarticular tubercles large, slightly ovoid, subarticular tubercle on Finger I larger than all others. Terminal discs expanded about 1.3 times on Finger I, discs expanded about 1.5 times on fingers II and III, and 1.6 times the width of the distal end of the adjacent phalanx on finger IV; round, paired dorsal scutes on each disc.
Hind limbs moderately robust, tibia length 48.73% SVL. Outer tarsal fold not evident, inner tarsal fold well developed, extending from mid-point of tarsus and connected to inner metatarsal tubercle. Inner metatarsal tubercle elliptical, raised about two times the size of bluntly rounded conical outer metatarsal tubercle. Toes basally webbed between toes II and III and between toes III and IV; webbing between toes II and III barely evident; webbing between toes III and IV more developed but not extending to proximal edge of first subarticular tubercle on either toe III or toe IV. Webbing absent between other toes, lateral fringes absent on all toes. Subarticular tubercles small, round, largest on toes I and II; first subarticular tubercle present but small on toe IV; supernumerary tubercles absent. Terminal discs of toes I, III, and IV expanded approximately 1.6 times the width of the distal end of the adjacent phalanx, disc on toe II expanded 1.8 times, and disc of toe V 1.5 times wider than distal end of adjacent phalanx.
Skin of venter smooth, skin of dorsum posteriorly tubercular, smooth anteriorly, but with scattered low tubercles also evident on the lateral margins of the dorsum anteriorly (with skin smooth medially except for low rounded granules posteriorly, clustered just anterior to the anus). Anal sheath short; anal opening directed postero-ventrally at upper level of thighs, cloacal tubercles absent. Tongue elongated, free posteriorly for almost 3/4 of its length, not notched posteriorly; median lingual process absent. Choanae round, widely separated; vomerine odontophores absent. Vocal slits present.
Coloration in life. Dorsum of head and body brown to dark brown to charcoal with few, faintly evident, scattered irregular brown to black spots. Flanks brown to black, with a cream oblique lateral stripe extending from the groin anteriorly to about level of mid-body. The black flank surfaces end in a distinct linear border with the white ventral surface, and the mustard-yellow pigment around the arm and leg insertions; aside from this sharply delineated border, no distinct separate pale ventrolateral stripe is evident in life or after preservation. The flanks are dark brown-gray anteriorly and light gray washed with cream posteriorly. Lateral surfaces of head (from below canthus rostralis to the ventral margin of the eye) dark brown to black; upper lip (from ventral margin of eye to mouth) cream, with a faint wash of pale gray pigment. Dorsal surfaces of legs light brown with diffuse dark brown markings consisting of 3 transverse dark brown bands on dorsal surfaces of thighs, with a thin longitudinal stripe of the same color running posteriorly the length of each thigh; shanks with 4 transverse bands of the same color combination as bands on thighs; ankles also with dark brown transverse bands over brown dorsally. Ventral, and proximal antero-ventral surfaces of thighs bright mustard-yellow (remaining antero-ventral surfaces of thighs light brown); this mustard-yellow pigment also extends across the groin, and posterior belly. Light mustard-yellow paracloacal marks present. Ventral and antero-ventral surfaces of shanks also are bright yellow, yellow extending onto the anterior surface of the ankle and fading along the proximo-dorsal surface of the foot. Dorsal and anterior surfaces of arms light brown with dark brown transverse bands and irregular marks; ventral surfaces of arms and hands gray. Proximal 1/2 of the posterior and postero-ventral surfaces of upper arms bright mustard-yellow (same color as on thighs), extending in an arc around the dorsal, posterior, and ventral circumference of each arm). This arc of mustard-yellow pigment is narrowest on that portion of the flanks of the body immediately dorsal to the arm insertion (where it ends at a sharply-defined border with the black lateral surfaces), and then broadens posteriorly (forming a crescent-shaped mustard-yellow mark also defining a crescent-shaped border with the black lateral surfaces), and ventrally (the ventral portion of this half-circle is the largest, with mustard-yellow pigment extending to a radius around the postero-ventral area of the arm insertion to a width of approximately 1/2 the length of upper arm).The iris was gold to copper/brown, with scattered small irregular black markings.
Coloration in preservative. Overall, the dark pigments on the dorsal and lateral surfaces of the body and limbs appear darker and less bright than in life, especially for pattern features that are narrow or small. All major features of pattern (pale or dark bands and stripes, ground color) remain nearly as distinct as in life despite this darkening. The white ventral ground color has faded to a dull cream, but darker pigments remain clearly visible aside from some fading, and all areas that were bright mustard-yellow in life have lost all trace of pigment, leaving only the underlying ground color (dull cream in ethanol).
Variation. Measurements of three adult female paratypes, and the ranges and means of measurements of the four adult male paratypes are presented in Table 3. We excluded from Table 3 two adult female paratypes that were lab-raised. The type series includes three subadult females, and ten unsexed juveniles; the three sub-adult females ( INABIO 3649 , INABIO 3785 , and KU 327677) are 20.1, 22.0 and 22.9 mm SVL , respectively. The ten unsexed juveniles measure between 12.6–15.2 mm SVL.
In some individuals, the patches and spots of dorsum are more evident over the background brown color. The posterior flanks vary from cream to gray with the pattern from irregularly spotted to reticulated, gradually fading into the pale ventral pattern. In some specimens, the longitudinal stripe running the length of each thigh is bordered posteriorly by a lighter stripe. Bright mustard-yellow markings in the groin and upper arm areas were clearly visible in life in all specimens. The paracloacal mustard-yellow marks are also present varying from light (holotype) to more contrasting (e.g. CJ 7765, Fig. 5A View FIGURE 5 ). In all specimens, the bright mustard-yellow appeared to be a separate outer layer of pigment overlying the ground color of white, which faded very rapidly following preservation (within 24–48 hrs.), leaving only the white visible.
Black lip stripes occur along the lips. One in the upper lip and another along the dorsal margin of the lower lip ( LLS). They occur in all specimens (with the line complete around the entire lower lip, and visible in preservative), but considerable variation is evident in the type series. In life, the LLS vary from a sinuous faint gray line visible only on close inspection of the dorsal margin of the lower lip, to a prominent straight black line that is obvious even from some distance ( Fig. 4E View FIGURE 4 ). The LLS of paratype QCAZ 14699 View Materials was more prominent in life than any other specimen, as is the LLS in preservative of adult female MCZ A-14025. The LLS is generally darker and more obvious in larger individuals. Among the remaining adult and sub-adult preserved specimens, the LLS is clearly visible under magnification, with no indication of sexual dimorphism .
The pale oblique lateral stripe (OLS) of Leucostethus bilsa (among adults and sub-adults) varies in length and continuity, with some specimens exhibiting a complete OLS from groin to eye, and others exhibiting a faded, broken, or incomplete OLS ( Figs. 3A, B View FIGURE 3 , 4 View FIGURE 4 A–D). It fades to gray-brown anteriorly in some individuals; for example, in KU 327677 (sub-adult female) and KU 327679 (adult male) extending only from the groin to level of mid-body in the flank. The OLS is not present in three juveniles, but clearly evident in sub-adult specimens.
There is sexual dimorphism in ventral coloration pattern. In the adult male holotype CJ 8311 and all four-adult male paratypes, the anterior ventral surfaces (chin and throat) are spotted gray over cream to white ground color (in life and after preservation), with the dark pigment forming a reticulated pattern in some individuals; the dark spotting also continues on venter in both of the adult males found carrying larvae ( INABIO 3632 and KU 327679), extending to a point approximately one-half of the distance between arms and thighs. In both sub-adult and adult females, the anterior ventral surfaces (chin and throat) are cream to white with a faint and diffuse pattern of pale brown to gray flecks (in life and after preservation); in some individuals, these flecks coalesce to form a reticulated pattern, but this is not pronounced and can be readily seen only under magnification, and the entire abdomen is unmarked cream to white .
No pronounced differences in color pattern among the three immature females and the six wild-caught adult specimens (2 females, 4 males) were noted, suggesting that ontogenetic color pattern changes are complete once the frogs reach ~ 20 mm SVL (although no sub-adult males are included in the type series). The three post-metamorphic unsexed juveniles were all nearly uniform brown or dark brown in life both dorsally and laterally, with oblique lateral stripes only very faintly evident under magnification. Despite the differences in dorsal, lateral, and oblique lateral stripe color as compared to sub-adults and adults, all three juveniles also exhibited obvious bright mustard-yellow flash marks in the axillary and groin regions. One noteworthy observation among these three juveniles pertains to their relative sizes in comparison to the respective lengths of their tail remnants; the two smallest juveniles ( QCAZ 20137 View Materials and KU 327678) measure 12.8 and 13.4 mm SVL, with tail remnants of 1.7 and 0.7 mm, respectively. In contrast, the largest juvenile ( QCAZ 14701 View Materials ) measures 14.0 mm SVL, yet has a tail remnant measuring 5.5 mm in length. There is no evidence of any ontogenetic change in ventral pattern .
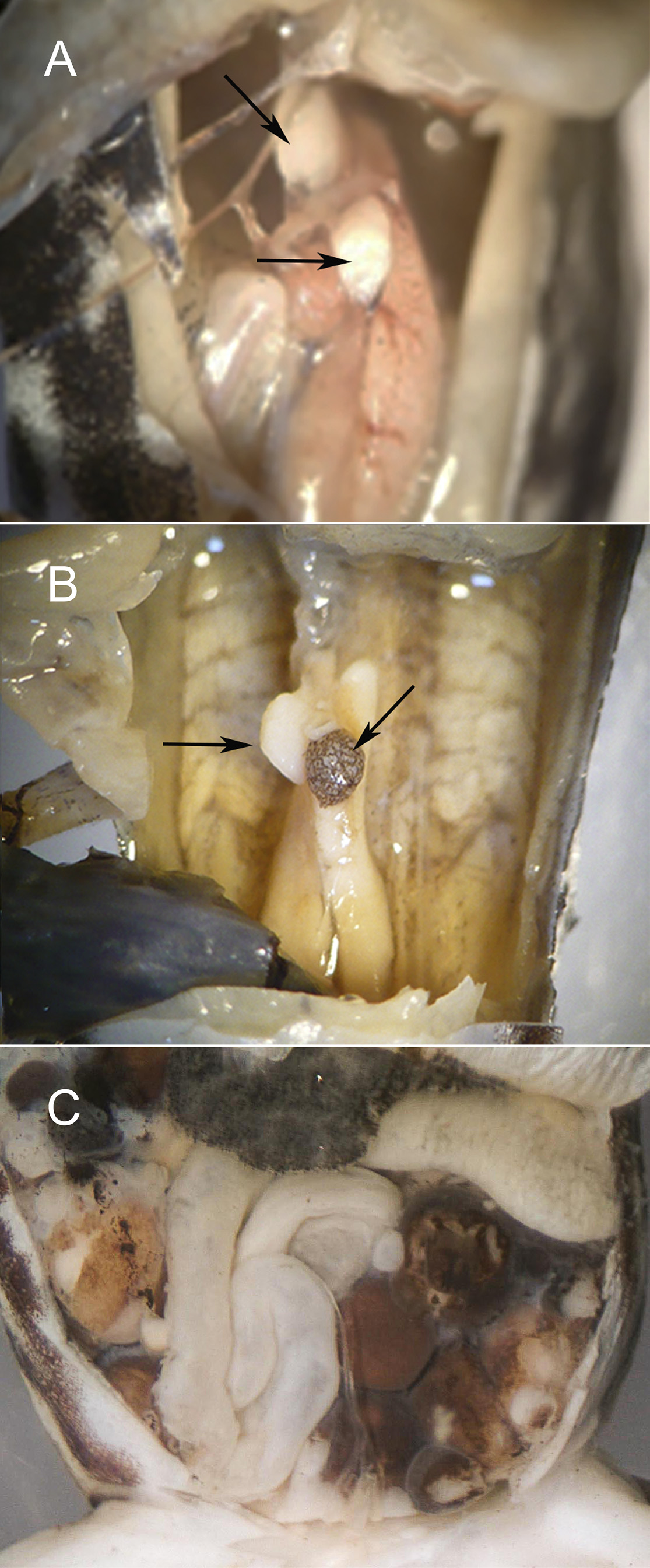
The color of the testis varies intraspecifically and intraindividually. Two adult males of C. bilsa ( QCAZ 14700 View Materials and INABIO 3786 ) each have one white (right) and one brown (left) testis (see Fig. 7 View FIGURE 7 ). However, both testes are white in the adult male holotype CJ 8311 , and one adult male, INABIO 3632 ( Fig. 7A View FIGURE 7 ), while both testes are brown in another ( KU 327679). Oocytes in advanced stages are brown ( Fig. 7C View FIGURE 7 ) .
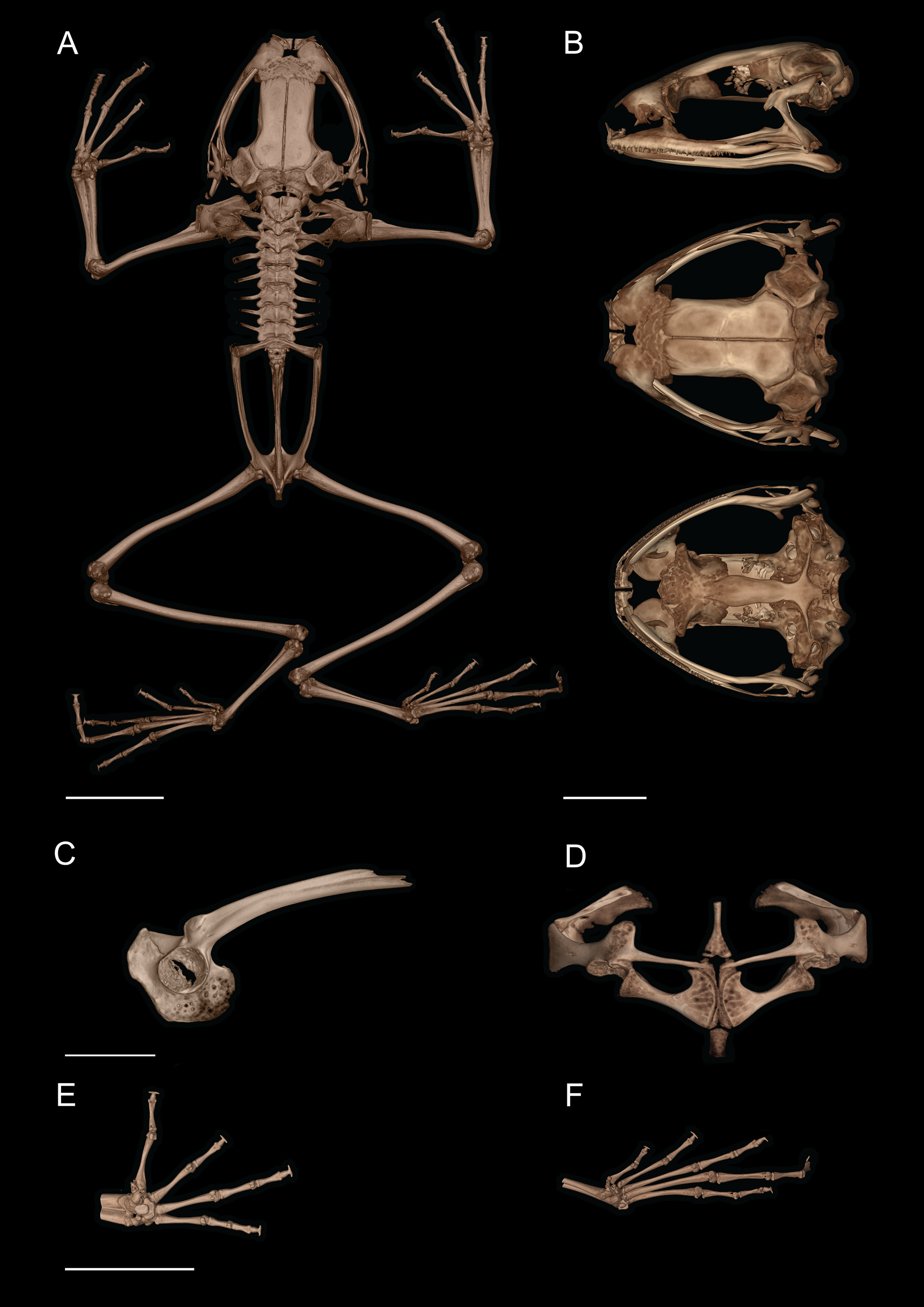
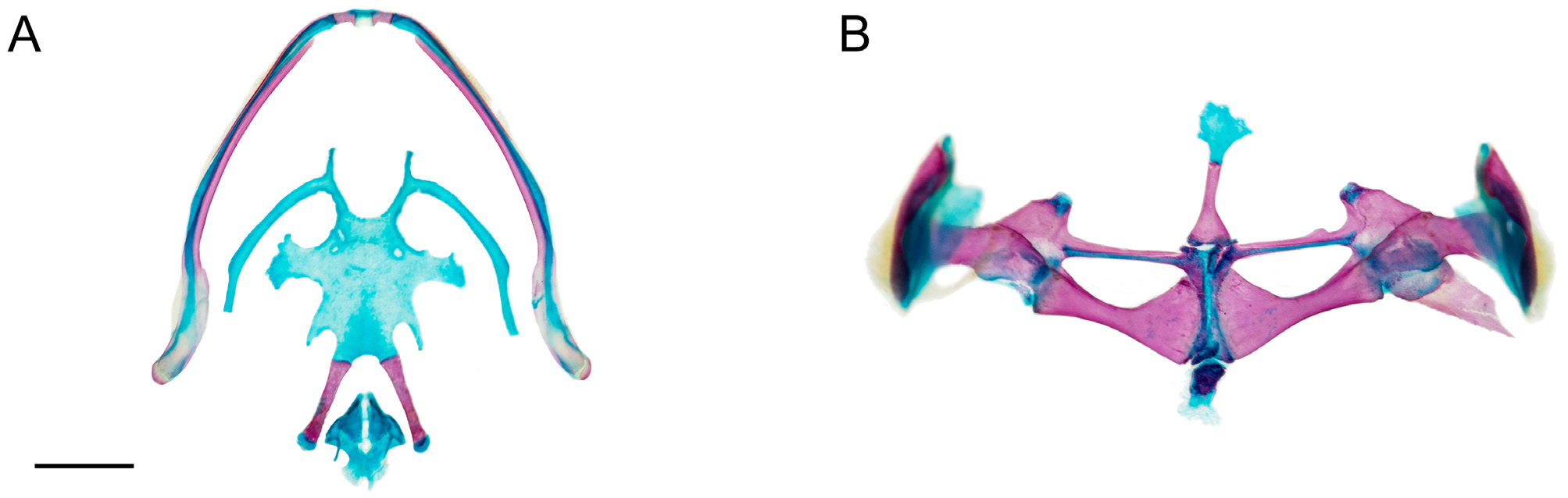
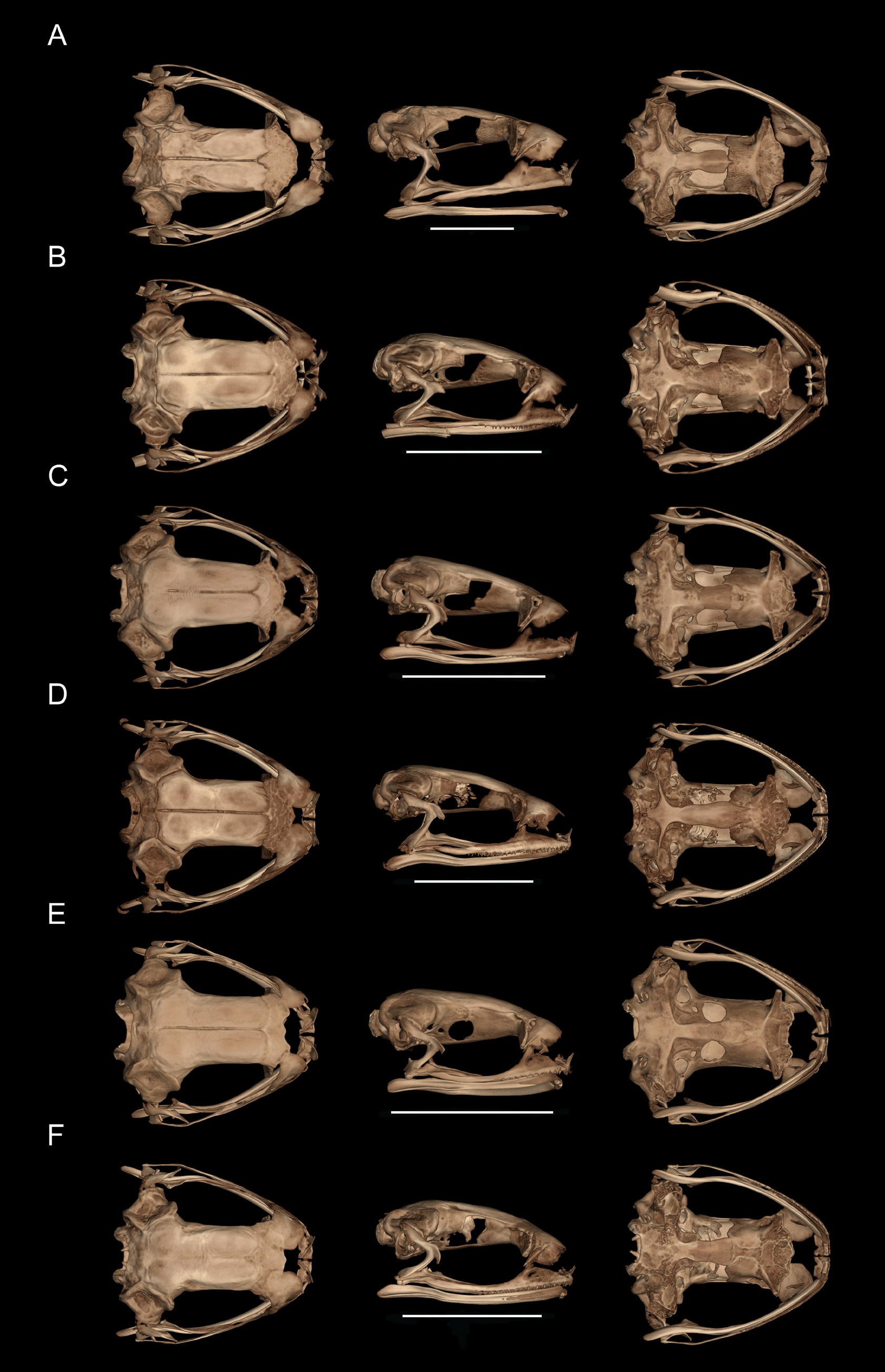
Osteology. General osteological features of the male holotype ( CJ 8311 ) are depicted in Fig. 8 View FIGURE 8 , and the hyobranchium and pectoral girdle of a paratype ( CJ 7775 , female) are shown in Fig. 9 View FIGURE 9 .
Cranium. The skull is longer than wide, about half as tall as long (skull length: 8.9 mm; skull width: 8.2 mm; skull height: 4.6 mm), lacks dermal ornamentation, but has rugose surfaces on some bones. The neurocranium is moderately ossified. The septomaxilla is small and round. The nasals are subcircular, bearing an acuminate posterolateral maxillary flange, but do not articulate with the pars facialis of the maxillary. The nasals are widely separated medially. The ossified sphenethmoid slightly underlies the posterior margins of nasals. The frontoparietals overlie the anteromedial prootic posterolaterally; the orbital edges of the frontoparietal are straight. The frontoparietals lack medial articulation. The crista parotica is well developed, long, and narrow; the occipital condyles are widely separated. The squamosal is moderately developed; the otic ramus articulates with the cartilaginous lateral margin of the crista parotica. The pterygoid is moderately developed, the medial ramus is not in contact with the ossified prootic; the anterior ramus extends to the anterior orbit and is in broad contact with the cartilaginous pterygoid process of the palatoquadrate adjacent to the pars dentalis of the maxilla. The quadratojugal is slender and articulates with the posterior end of the maxilla. The maxilla and premaxilla bear mostly monocuspid teeth, with a maximum number of 5 teeth cusps in the premaxillae and 32 in the maxillae. The maxillae are slender, lacking a postorbital process; the preorbital process is moderately well developed; the pars palatina of maxilla is narrow. The alary process of the premaxilla is narrow and curved posterolaterally; the pars palatina of the premaxilla is well-developed and broadly “U”-shaped in ventral aspect. The mentomeckelian bone is slightly constricted, about equal in width at medial and lateral ends. The angulosplenial extends posterior to the jaw articulation. The vomer is small, edentate, and bears a small pre- and a large postchoanal process bordering the anterior and lateral margins of the choana, respectively. The neopalatine is absent. The planum antorbitale is cartilaginous. The parasphenoid is “T”-shaped and lacks dermal modifications; the alae are acuminate, directed slightly posterolaterally; the cultriform process is about twice as wide as transverse length of alae; the anterior end of the cultriform processes bears large bifid projections and extends to the anterior region of the orbit. The pars media plectri of columella is slender. The tympanic annulus is sickle-shaped, with the open side upper. The tympanic region bears a cartilaginous round plate.
Hyolaryngeal apparatus. The hyoid plate is longer than wide (measured at midlines) and bears three holes at its anterior portion. The hyale are long, slender, angling posterolaterally to the otic region, wider in proximal than distal half, and bearing a slightly expanded region at the midline. The hyale bears one anterior process; it is straight and its distal end is rounded. The hyoglossal sinus is circumscribed “U–V”-shaped. The alary processes are basally stalked and relatively large. The posterolateral processes are slender and straight. The posteromedial processes are ossified. The cricoid ring is complete.
Vertebral column. There are eight procoelus presacral vertebrae with nonimbricate neural arches; the cervical cotyles are widely separated, the cotyles are stalked (cervical cotylar arrangement Type 1 of Lynch 1971). The neural spines decrease in height progressively from anterior to posterior and have a keeled upper border. The transverse processes of Presacral III are long; the transverse processes of the posterior presacral vertebra are short; and the transverse processes of Presacral II are the shortest. The transverse processes on Presacrals II–VIII bear cartilaginous distal ends, which are calcified in Presacrals III–VIII. The transverse processes of Presacrals II, VII and VIII are directed anteriorly, and those of Presacrals III–VI are directed nearly posteriorly. The sacral diapophyses are not dilated and directed slightly posteriorly. The sacrococcygeal articulation is bicondylar. The coccyx has a narrow dorsal ridge extending the entire length of coccyx; the coccyx is moderately wide anteriorly; the transverse processes of the coccyx are absent .
Pectoral girdle. The pectoral girdle is firmisternal; an omosternum is present. The style of the omosternum is ossified. The distal end of the style bears a short, broad cartilage. The xiphisternum is rectangular and basally calcified. The clavicles are oriented horizontally, forming an L with the outer lateral extreme. The coracoids are moderately robust, oriented so that the sternal end lays posteromedial to the glenoid end. The medial margin of the pectoral fenestra is formed by the coracoid; the calcified epicoracoids are abutting one another, narrowly separated anteriorly. The procoracoid cartilage is present, of uniform depth throughout transverse width. The cleithrum is distally bifurcate. The suprascapula lacks calcification in the margins.
Pelvic girdle. The pelvic girdle is “V”-shaped in dorsal view; the ilial shafts extend anteriorly slightly anterior to the sacral articulation. The dorsal crest of the ilium is present. The preacetabular zone is not expanded ventrally; the postacetabular zone is not expanded. They are juxtaposed at ventral midline by a cartilage.
Appendages. The deltoid crests on the humerus are prominent. The phalangeal formula of the hand is 2-2-3-3. The prepollex is composed of two elements, with the basal element calcified. The distal transverse processes of the terminal phalanges on the fingers and toes are twice the width of the terminal phalange at midlength. The astragalus and calcaneum are fused proximally and distally. The phalangeal formula of the foot is 2-2-3-4-3. The terminal phalanges are “T”shaped. The prehallux is composed of 2–3 cartilaginous elements; the basal element is calcified. There are three ossified tarsal bones and two tarsal sesamoid bones. There are one or two large sesamoid bones at the joints of phalanges of fingers, smaller and single ones on the toes.
Vocalization. A chorus of three or four individuals was heard calling from the forest floor in a relatively flat area of forest at the base of a steep slope beside the Green trail, ~ 1.2 km SW of the RBB main station complex on the morning of 5 January 2004. These individuals were calling within a relatively small area, spaced about ten meters apart. Typically, when one frog commenced calling, the other frogs would then start calling; between periods of calling continuing for only a few minutes, there would be long periods of silence. The individual recorded was perched in a prominent position on a dead leaf, raised a few centimeters above the rest of the leaf litter on the forest floor. Only a few individuals were heard calling elsewhere at Bilsa from 3–10 January 2004, and so L. bilsa was presumed to be uncommon at that time, but during the rainy season in February of 2007, calling males were more commonly heard in many areas, suggesting that L. bilsa may be fairly abundant at the RBB.
The call is a series of peep notes ( Fig. 10 View FIGURE 10 ) with a series of 65–88 peeping notes repeated at 427–739 ms intervals (mean = 504, SD = 92, N = 20), the entire sequence lasting from 46.8– 59.3 s in duration. Two inter-call periods are 25 and 68 s. Overall note repetition rate was 1.35 notes per second, or 81.2 notes per minute. The notes were produced at regularly spaced intervals, (inter-note interval being 431–471 ms, mean = 443, SD = 16); however, the inter-note interval was shorter between some note pairs resulting in a double peep being heard. Only 12 double peeps were present in the entire sample of 237 notes and were never emitted more than three times in succession. In double peeps the inter-note interval was shorter (336–416 ms, mean = 363 ms N = 3), and between the pairs of double notes was longer (660–666 ms, mean = 662 ms, N = 3). Each peep was a weakly pulsatile frequency modulated note (rising from lower to higher) with a duration of 127–182 ms (mean = 153, SD = 17). The dominant frequency arose from 3310–3398Hz (mean = 3340, SD = 28.9) to 3660–3722 Hz (mean = 3691, SD = 25.4). The fundamental frequency was about 1660 Hz rising to 1820 Hz. Four higher harmonics were present, their starting frequencies being about 4940, 6620, 8360, and 9890 Hz respectively.
Distribution and natural history. Leucostethus bilsa is only known from RBB, Provincia Esmeraldas, borderline between Esmeraldas and Manabí provinces, in the Chocoan region of Pacific northern coastal Ecuador, from elevations between 426 and 515 m, in an area of extent of occurrence of only about 6 km 2.
This diurnal, riparian species inhabits primary or older secondary forest in the Evergreen Forest of Lowlands of Ecuatorial Chocó and the Evergreen Seasonal Montane Foothill Forest of Cordillera Costera of Chocó, where the average annual rainfall is 1759–2566 mm and the average annual temperature is 22.2–25.4 ºC. All specimens of Leucostethus bilsa were found moving on surface leaf litter during the day along streams and adjacent slopes. The holotype CJ 8311 was collected along the Brown trail, approximately 1.3 km NE from the main station complex. Other specimens included as type series, here described, were collected along the Green and Black trail routes. Two juvenile paratypes were collected between rocks and leaf litter in the margins of the Duchas stream ( Fig. 1B View FIGURE 1 ), close to a 10-meter waterfall. Other paratypes were collected in an old secondary forest along the White trail. One adult male ( INABIO 3632 , transporting larvae) was captured in primary forest on a large flat rock near the Aguacatal–Du-chas stream .
Other dendrobatid frogs occurring sympatrically at Bilsa include Epipedobates boulengeri ( Barbour 1905) , Hyloxalus awa and Oophaga sylvatica .
Three adult males were transporting larvae when found. Two adult males ( KU 327679: August, 2002, and INA- BIO 3632 : December, 2006) were observed carrying larvae when encountered, although for both males all larvae were lost during capture; the former was found only ~2 meters from water in a streamside area strewn with rocks, and the latter was found on a rock, also very near a stream, carrying an estimated 5– 6 larvae (dark brown to black in color). A third male ( CJ 7213 ) was observed carrying 11 larvae ( Fig. 4F View FIGURE 4 ) on 2 August 2017, under a rock covered by leaf litter in the Duchas stream. The latter tadpoles were preserved in RNAlater solution for molecular analyses. A lab-raised female had a maximum mass of 2.2 g. The juveniles (post-metamorphs) were found in December, February, March, May, and June. The presence of tadpoles and metamorphs from August to June is indicative that breeding occurs throughout the year .
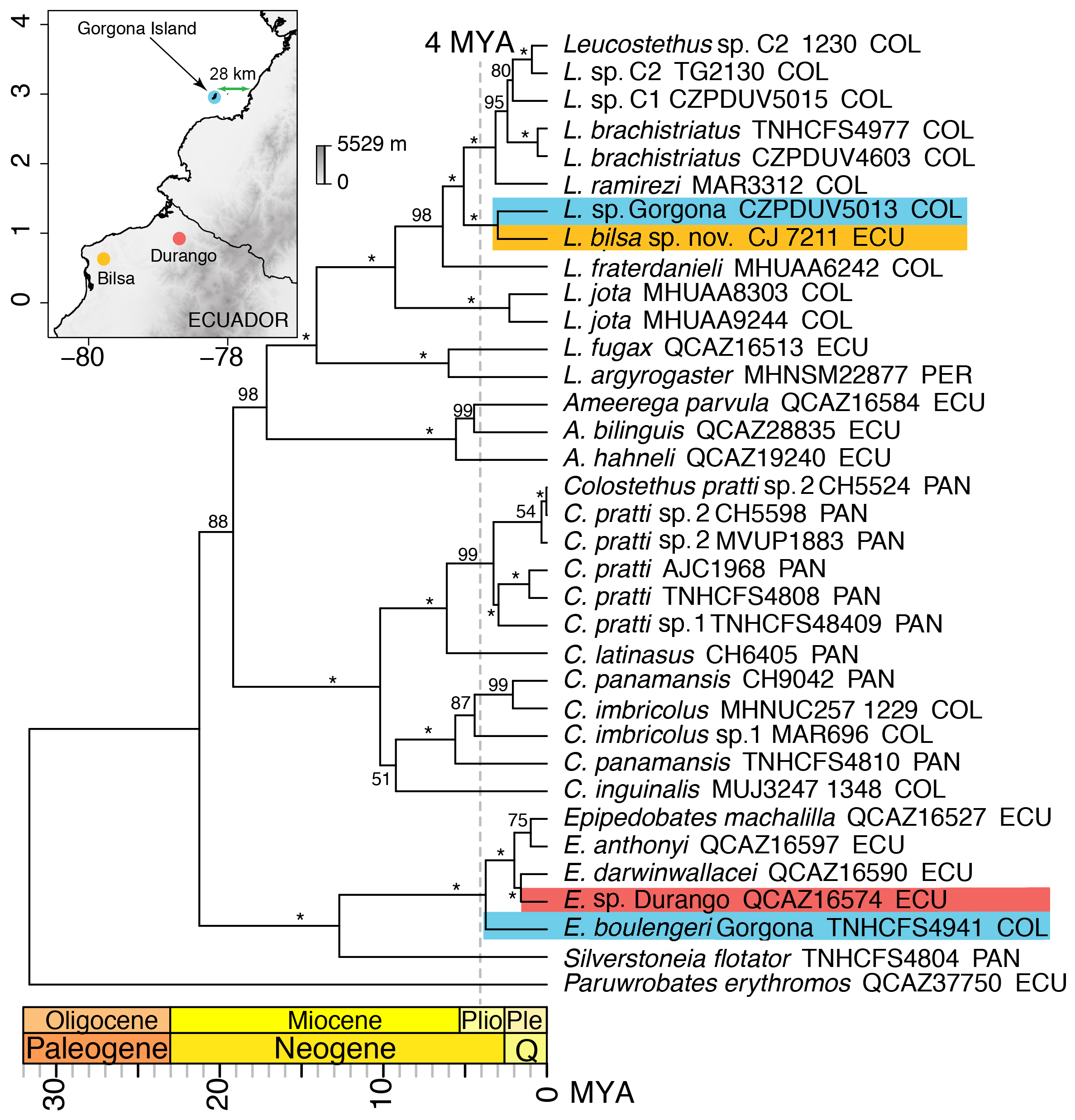
Phylogenetic analysis and systematics. The new species was found to be nested in Leucostethus ( Fig. 11 View FIGURE 11 ) and it is the sister taxon to specimens of an undescribed species (L. sp. Gorgona) from Gorgona Island, located 28 km off the southwest of the Colombian Pacific coast (e.g., CZPDUV5013–14). There are 6.25% (i.e., 146/2335 bp) differences for the section of 12S-16S mitochondrial fragment, between L. bilsa and L. sp. from Gorgona. With the species described here, the taxa included in Leucostethus with molecular data are Amazonian species (i.e., L. argyrogaster and L. fugax ), Chocoan and Northwestern Andean species (i.e., L. brachistriatus , L. fraterdanieli , L. jota , L. ramirezi , and L. bilsa ) and several populations of undescribed species (i.e., L. sp. C1, L. sp. C2 and L. sp. Gorgona). Overall, our estimated phylogenetic tree agrees with that of Marin et al. (2018) and supports their conclusions about Leucostethus fraterdanieli as a complex of species nested within the recently named Leucostethus ( Grant et al., 2017) . This genus is well-supported (100 ML bootstrap) and sister to Ameerega . The clade Ameerega + Leucostethus is well-supported (98 ML bootstrap) and sister to Colostethus as redefined by Marin et al. (2018). Finally, the clade that includes Ameerega + Leucostethus + Colostethus is also well-supported (88 ML bootstrap).







| BBS |
University of Suriname |
| MCZ |
Museum of Comparative Zoology |
| KU |
Biodiversity Institute, University of Kansas |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Leucostethus bilsa
| Vigle, Gregory O., Coloma, Luis A., Santos, Juan Carlos, Hernandez-Nieto, Sebastian, Ortega-Andrade, H. Mauricio, Paluh, Daniel J. & Read, Morley 2020 |
Colostethus
| Cope 1866 |