
Afropinnotheres Manning, 1993
|
publication ID |
https://doi.org/10.11646/zootaxa.3947.2.8 |
|
publication LSID |
lsid:zoobank.org:pub:B7F32AB1-3F28-467C-916F-A72EE3307D02 |
|
DOI |
https://doi.org/10.5281/zenodo.6100816 |
|
persistent identifier |
https://treatment.plazi.org/id/03BE87DA-1674-FFC1-FF15-FA38FD2B9ADF |
|
treatment provided by |
Plazi |
|
scientific name |
Afropinnotheres Manning, 1993 |
| status |
|
Genus Afropinnotheres Manning, 1993 View in CoL
Afropinnotheres Manning, 1993: 130 View in CoL .
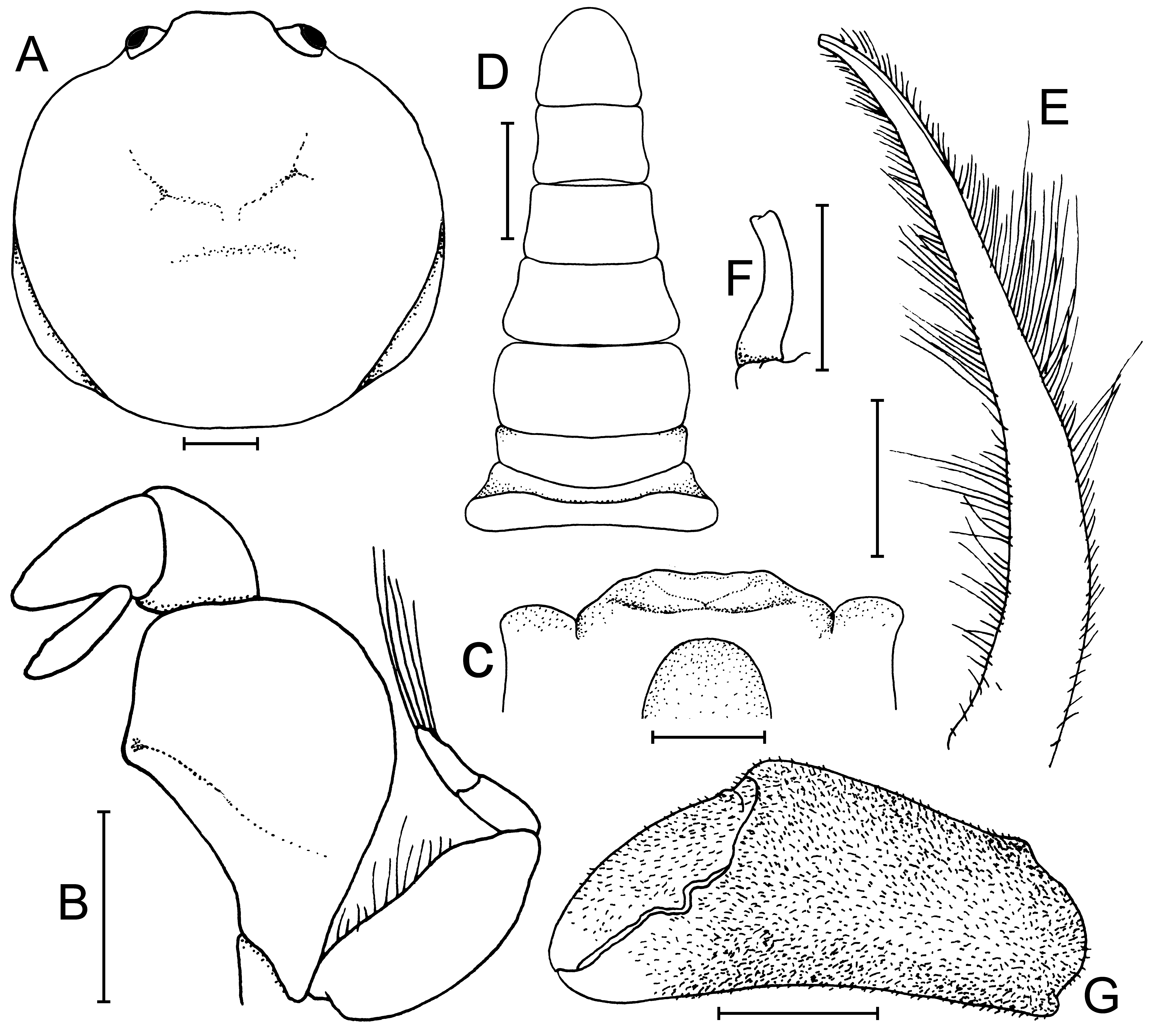
Remarks. Manning (1993) substantially changed the generic system for the species placed in Pinnotheres Bosc, 1802 , differentiating genera mainly on the basis of the structure of the MXP3 (notably the structure of the dactylus and its point of articulation with the propodus) and the proportions of their P2–P5 (notably the dactylus). Using these and characters of the carapace in some instances, over a dozen genera are now recognised (see Ng et al. 2008).
In reviewing the generic status of many of the Atlantic species, Manning (1993) established a new genus, Afropinnotheres , for species that share the following characters: a rounded to subovate carapace, the propodus of the P2–P5 are laterally flattened and appear expanded; the P2–P5 dactyli are subequal; propodus of the MXP3 is distinctly coniform and is shorter than the carpus; dactylus of the third MXP3 is elongated, extending beyond the tip of the propodus and articulates near the base of the propodus; all the male abdominal somites are free; and the male first gonopod is simple and slender. Afropinnotheres is similar to Pinnotheres Bosc, 1802 s. str., and Nepinnotheres Manning, 1993 , but in these genera, the propodus of the MXP3 is distinctly more elongated (much longer than the carpus), with the dactylus either long, reaching to or over-reaching the tip of the propodus and inserted at or near the middle or the proximal third of the propodus ( Nepinnotheres ), or the MXP3 is shorter, not reaching the tip of the propodus and inserted near the base of the propodus ( Pinnotheres s. str.). Another character shared by all Afropinnotheres species is the proportionately slender male abdomen. The known male abdomens of Pinnotheres and Nepinnotheres are proportionately broader, with the first few somites much wider, giving the structure a more triangular appearance (see Ahyong & Ng 2007).
Four species of Afropinnotheres have been described, all from western Africa: A. crosnieri Manning, 1993 ( Congo; type species); A. guinotae Manning, 1993 ( Angola) ; A. larissae (Machkevskiy, 1992) ( Senegal, Guinea, Ivory Coast); and A. monodi Manning, 1993 ( Morocco, Mauritania).
Ng & Manning (2003) subsequently transferred a number of species of holothurian-dwelling Pinnotheres to a new genus, Holotheres , members of which share carapace and ambulatory leg characteristics with Afropinnotheres . In addition to their host preference (all Afropinnotheres are from bivalves), however, Holotheres can be separated by the elongated propodus of the MXP3 (much longer than the carpus) and a more distinctly spatulate dactylus which is as long as or longer than the propodus; the male abdomen is more triangular in shape and the G1 is gently sinuous (cf. Ng & Manning 2003: fig. 1B, H, I).
Afropinnotheres View in CoL also closely resembles species of Pinnaxodes Heller, 1865 View in CoL , with regard to the carapace shape and symmetrical P2–P5. The MXP3 of Pinnaxodes View in CoL is superficially similar to that of Afropinnotheres View in CoL except that the propodus is elongated and much longer than the carpus (rather than short and conical in Afropinnotheres View in CoL ) and the more spatuliform dactylus is inserted on the proximal one-third or middle of the propodus (cf. Garth 1958: figs. 10D, 11B; Wells & Wells 1961: fig. 2F, G; Campos et al. 1998: fig. 2B, D; Takeda & Prince Masahito 2000: figs. 2C, E, G, 4C; Ng & Manning 1993: figs. 6B, F, 7A, B; De Melo & Boehs 2014: fig. 4). The male abdomen of Pinnaxodes View in CoL species is also relatively wider and more triangular in shape and the telson may be expanded such that it is wider than somite 6 (cf. Garth 1958: fig. 11A; Wells & Wells 1961: fig. 1F; Campos et al. 1998: fig. 3A, C; Takeda & Prince Masahito 2000: figs. 4B, 5L; De Melo & Boehs 2014: fig. 6).
There has been some discussion about the taxonomy of Pinnaxodes Heller, 1865 View in CoL , and Holothuriophilus Nauck, 1880 View in CoL . Although Takeda & Prince Masahito’s (2000) suggestions on these genera were provisional, De Melo & Boehs (2004) followed their arguments for transferring the Japanese Pinnaxodes mutuensis Sakai, 1939 View in CoL , to Holothuriophilus View in CoL and placed the South American P. tomentosus Ortmann, 1894 View in CoL , there as well. Ng & Manning (2003) showed that Pinnaxodes View in CoL and Holothuriophilus View in CoL are easily separated by the carapace shape (ovate in Pinnaxodes View in CoL , transversely subrectangular in Holothuriophilus View in CoL ) and form of the MXP3 (i.e. presence of a faint suture separating the ischium and merus in Pinnaxodes View in CoL ; completely fused ischium and merus in Holothuriophilus View in CoL ) (see also Ahyong & Ng 2007; Campos et al. 2012); and retained P. mutuensis View in CoL in Pinnaxodes View in CoL . Six species are now recognised in Pinnaxodes View in CoL : P. chilensis (H. Milne Edwards, 1837) View in CoL (= Pinnaxodes hirtipes Heller, 1865 View in CoL ) [ Chile to Per]; P. f l o r i d e ns i s Wells & Wells, 1961 [Florida, USA]; P. gigas Green, 1992 View in CoL [Sonora, Mexico]; P. m a j o r Ortmann, 1894 [ Japan and northeastern Russia]; P. m ut uensis Sakai, 1939 [ Japan and northeastern Russia]; P. tomentosus Ortmann, 1894 View in CoL [ Brazil] (Ng et al. 2008).
The matter of fusion between the MXP3 ischium and merus requires discussion. Of the many genera associated with Pinnotheres View in CoL s. lato in the past, only Pinnaxodes Heller, 1865 View in CoL , has these two structures separated by a suture, faint or otherwise (see Manning 1993: 128). Takeda & Prince Masahito (2000) noted that while it is present, they regard it as a less important character since it is not always apparent and can be hard to see. Ng & Manning (2003) noted that it was always present but may be shallow, with the suture more pronounced on the inner surface of the MXP3 and best observed under transmitted light. All known species of Pinnaxodes View in CoL possess this suture. The present new species from India, Afropinnotheres ratnakara View in CoL n. sp., also has a trace of this suture. There is a faint suture visible under transmitted light between the merus and ischium in the larger female specimens ( Fig. 2 View FIGURE 2 D), but it is not distinct even on the inner surface of the structure unlike Pinnaxodes View in CoL . This suture is even less distinct in the smaller males ( Fig. 3 View FIGURE 3 B) and may be absent. Manning (1993) did not report any trace of a suture for the four African species of Afropinnotheres View in CoL he studied, but he may have overlooked this feature; the MXP3 of the African Afropinnotheres View in CoL require restudy.
The discovery of the present new species from India is noteworthy because all other Afropinnotheres View in CoL species have previously been reported from the eastern Atlantic ( Manning 1993). The characters of Afropinnotheres ratnakara View in CoL n. sp., however, are distinct for the genus. As discussed earlier, there are affiliations with Pinnaxodes View in CoL as well, but this genus is also not known from the Indian Ocean. While it is possible that the presence of the new species in India is a consequence of introduction from the Atlantic, the fact remains that it may be the same species as specimens of Pinnotheres View in CoL sp. reported by Doflein (1904) and Tirmizi & Ghani (1996) from South Africa and Pakistan, respectively (see species discussion), many years earlier. Another possibility of course is that the host Perna perna may have been introduced from Africa to India decades or even centuries ago with the crab. This mussel has a rather disjunct distribution, occurring in East and South Africa as well as southwestern India; and the Perna in India may have come from South Africa, especially since Doflein’s (1904) Pinnotheres View in CoL sp. is probably conspecific with Afropinnotheres ratnakara View in CoL n. sp. In any case, the current evidence suggests that Afropinnotheres View in CoL is naturally present in the western Indian Ocean.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Brachyura |
|
Family |
Afropinnotheres Manning, 1993
| Ng, Peter K. L. & Kumar, Appukuttannair Biju 2015 |
Afropinnotheres
| Manning 1993: 130 |