
Euglossa dilemma, 2011
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2011.00740.x |
|
persistent identifier |
https://treatment.plazi.org/id/03BF8780-6736-FFFF-9BF0-FD58FC16144F |
|
treatment provided by |
Valdenar |
|
scientific name |
Euglossa dilemma |
| status |
sp. nov. |
EUGLOSSA View in CoL ( EUGLOSSA ) DILEMMA BEMBÉ & ELTZ , SP. NOV.
The chemical, molecular, and morphological dichotomies described above require a re-evaluation of the taxonomic status of what was previously considered E. viridissima Friese. The male lectotype of that taxon at the Natural History Museum in Vienna has two mandibular teeth (see below) and, unambiguously, belongs to the less common, predominantly bidentate lineage. As a consequence this lineage has priority over the name E. viridissima Friese , and we describe the more common tridentate lineage as a novel species.
Material and methods: The studied specimens belong to the entomological collections of the Zoologische Staatssammlung München ( ZSM), Germany, the Smithonian Institution (SI) in Washington, DC, USA, the Natural History Museum of Kansas (Snow Entomological Collections, SEC), the Collection of Thomas Eltz ( CTE), Bochum, Germany, and to the Collections of G. Gerlach ( CGG), Munich, Germany. The type series comprises 255 male specimens. Euglossa dilemma has been compared with the male lectotype of E. viridissima from the Naturhistorisches Museum Wien, Austria. The lectotype, which is also referred to by Moure (1970), is dirty, and missing the right antenna above the scape, left hindleg, and left forewing. Its right mandible, which bears two pointed teeth, is clearly visible. It has tree labels: ‘Bilimek Mexico 1871 // Euglossa § viridissima det. Friese 1898 // LECTOTYPE viridissima Friese, J. S. Moure 1958 ’. In the following, terga and sterna are referred to as T 1, T 2, T 3, etc., and S1, S2, S3, etc. Integument and setae coloration were described by eye using a Leica MZ 6 dissecting microscope.
Diagnosis: Males of E. dilemma can be distinguished from those of E. viridissima by two main morphological features: males of E. dilemma have three man-
angle; in E. viridissima the hind tibia is slightly more obtuse and rounded ( Fig. 5 View Figure 5 ).
dibular teeth, with the intermediate tooth placed approximately at half distance between the other two teeth ( Fig. 3B View Figure 3 ), and their hind tibia is less inflated, with its distal/posterior edge pointing in a more acute
DESCRIPTION
Male: Colour and vestiture. Whole bee metallic green, darkest green on front of clypeus (seldom with bluish hue) and hind legs, lighter green on face near antennal scape and behind the ocelli, on propodeum, pleurae, and on T7. Violet stripe in front of velvet area on mid tibia. Clypeus and front of head with white setae, on top of head mixed with black setae; scutum and scutellum dorsally covered with a mix of many white and some black setae, end of scutellum only white; terga with tiny white setae, on top of T2–T5 some black setae; sterna with white setae, centre of S2 with two large joining cushions of dense brownish hairs; wings transparent, covered with short black setae, wing venation dark brown. Jugal comb at base of hind wing with 16 blades.
Head. Width 4.6 mm, height from labrum to front 2.6 mm. Mandibles with three teeth, with the intermediate tooth placed approximately at half distance between the other teeth ( Fig. 3B View Figure 3 ). Tongue in repose reaching S2. Front of clypeus densely punctuated, with a complete medial rigde; ivory paraocular marks well developed, reaching the malar area; forward side of antennal scape with white stripe occupying two-thirds of its length. Labrum white with a medial ridge and two oval transparent windows.
Thorax and metasoma. Total body length 10.5– 11.5 mm; anterior wing 9.2 mm; scutum 2.9 mm wide (intertegular distance) and 2.7 mm long; scutellum 2.7 mm wide and 1.3 mm long. Punctuation on scutum dense and regular, all punctures have nearly the same size. Scutellum rounded on posterior margin. Punctuation on scutellum not as dense as on scutum. Anterior rim of scutellum with small punctures, at the centre and towards the posterior margin with larger punctures as scutum, nearly as many micropunctures as punctures. Median depression covering about two-thirds of scutellum, depression without punctures or with small punctures only. Punctuation on metasoma. Anterior half of T1 with sparse, large punctures; posterior half with dense, small punctures; T2–T4 with dense, smaller punctures as scutum; punctures in T5–T7 larger, and sparser.
Legs. Mid tibia, anterior tuft smaller than posterior tuft, triangular or comma-like. Posterior tuft oval or with drop shape, very close to anterior tuft. Generally, mid-tibial tufts indistinguishable in size and shape from those of E. viridissima . Velvet area dense on anterior end, sparse and incomplete on posterior end. Measurements are given as mean ± SD. Results of an ANOVA (d.f. = 1, 243) are also given and significant differences (P <0.01) are highlighted in bold letters and numerals.
Hind tibia triangular, 3.3 mm long and 2.6 mm wide. Basal third densely punctured. The distal two-thirds sparsely punctured; distance between punctures one or two diameters of punctures; punctures long, as many micropunctures as punctures. Hind tibia not as obtuse and distal/posterior edge less rounded as in E. viridissima ( Fig. 5 View Figure 5 ). Post-glandular area fringed with medium-sized hairs.
Female: Similar to male in size, coloration, punctuation, and vestiture. Paraocular marks, white stripe on antennal scape, or tufts on legs and S2 absent. Tongue in repose reaching S2. Mandible tridentate, distal and middle tooth ending in a point or rounded, basal tooth flat and broader than other two teeth. Scutellar tuft one-third to nearly half scutellar length. Hind tibia rounded with strong black or white setae, shiny but covered with micropunctures.
Type locality: Holotype collected at Xmatkuil near Mérida, Yucatán, Mexico (20°52′11.47″N, 89°37′ 10.03″W), April 2006, coll. Thomas Eltz. GoogleMaps
Distribution: From Mexico to Costa Rica. Recently introduced into southern Florida, USA (see also Geographical distribution below).
Etymology: The species epithet refers to the dilemma of having to describe a taxon as a novel species although this taxon had been widely considered as an existing species (dilemma is a noun in apposition).
Type material: HOLOTYPE. Male , with the following label data: ‘ Mérida (Xmatkuil) Yucatán, Mexico April 2006, coll. Thomas Eltz // Holotypus, male, Euglossa dilemma, Bembé & Eltz, 2010 ’ ( ZSM) . PARATYPES. 254 males, with the following labels data: 1 § ‘ MEXICO. Quintana Roo, 12 km NW. Reforma, 14 October 1986, C. D. Michener et al.’ ( SEC); 1 § ‘ MEXICO. S. Luis Potosí, Tlamaya Falls nr. Xilitla , 600 m, 1-IX-1991, D. Yanega’ ( SEC); 1 § ‘ MEXICO. Chiapas, 5 km S Cacahoatan, 340 m, 14°57′N, 92°09′W, 18-IV-1993, E. Tovar’ ( SEC); 2 §§ ‘ MEXICO. Veracruz, 5km S Boca del Rio. .. Jan. 2, 1982, B. H. Smith’ ( SEC); 2 §§ ‘ MEXICO. Veracruz, “Dunas Costadas”... Jan. 2, 1982, E. M. May’ ( SEC); 10 §§ ‘GG-1739 – 1748, Mexico, 8/99, Quintana Roo, Las Panteras (bosque alto), coll. G. Gerlach, Köder: p-Dimethoxybenzen’ ( CGG); 1 § ‘ Mexico, Bacalar, 18°45′n. B., 88°40′w. L., 10 m ü NN, 10. 1. 1998, leg. B. Bembé’ ( ZSM); 1 § ‘ MEXICO, Veracruz’ ‘ XI-1963, NLH Krauss’ (SI); 23 §§ ‘Merida (Xmatkuil) Yucatán, Mexico, 13.–27. 10. 2006, coll. Thomas Eltz’ (6: CTE, 15: ZSM, 1: Collection Linz, Austria, 1: Universidade Federal de Minas Gerais , Belo Horizonte, Brazil); 1 § ‘Mayapán, Yucatán, Mexico, 13.–27. 10. 2006, coll. Thomas Eltz’ ( CTE); 7 §§ ‘Xmatkuil, Yucatán, Mexico, at chemical bait, September 2009, coll. Thomas Eltz’ (1: CTE, 6: ZSM); 4 §§ ‘Merida (Xmatkuil) Yucatán, Mexico, April 2006, coll. Thomas Eltz’ (1: CTE, 3: ZSM); 10 §§ ‘Chiquilá, Quintana Roo, Mexico, at chemical bait, September 2009, coll. Thomas Eltz’ (4: CTE, 6: ZSM); 3 §§ ‘Chetumal, Quintana Roo, Mexico, at chemical bait, September 2009, coll. Thomas Eltz’ ( CTE); 1 § ‘Atasta, Campeche, Mexico, at chemical bait, September 2009, coll. Thomas Eltz’ ( CTE); 3 §§ ‘ Mexico, Veracruz, UNAM /Catemaco, VIII 92, coll. Thomas Eltz’ (2: CTE, 1: ZSM); 1 § ‘ EL SALVADOR, San Salvador, 23. May 1958, OL Cartwright’ (SI); 1 § ‘...?’ (SI); 1 § ‘ Qualan Quat, J E 4. 22. 1912 ’ (SI); 2 §§ ‘Belize, Sartaneja, Shipstern Reserve , 19°30′n. B., 88°20′w. L., 10 m ü NN, 12. 1. 1998, leg. B. Bembé’ ( ZSM); 2 §§ ‘ Guatemala, Santa Rosa, Monterrico, 25. VIII. 92, coll. Thomas Eltz’ (1: CTE, 1: ZSM); 2 §§ ‘GG-1459 – 1460, Guatemala, 4/98, San Lucas Toliman, Finca Faser, coll. P. Josefidou, Köder Eugenol’ ( CGG); 2 §§ ‘ Honduras, Utila, 16°15′n. B., 86°50′w. L., 20 m ü NN, 15. 2. 1998, leg. B. Bembé’ (1: ZSM, 1: CTE); 1 § ‘Honduras, Copan Ruinas, 14°35′n. B., 89°08′w. L., 3. 2. 1998, leg. B. Bembé’ ( ZSM); 1 § ‘ Honduras, la Ceiba, 4. 2003, leg. H. Heider’ ( ZSM); 1 § ‘ HONDURAS: Atlantida, Lancetilla Bot. Grd., Tela, 10 m, 22 VI 1994, 15°64′N, 87°27′W, Ashe, Brooks, ex: methyl salicylate’ ( SEC); 1 § ‘ HONDURAS: Atlantida, La Ceiba, 15 km W, 175 m, June/ July 1996, R. Lehmann, ex: methyl salicylate’ ( SEC); 3 § ‘GG-1125 – 1127, Nicaragua, 12/96, L. de Nicaragua, Insel Ometepe, coll. Anette Müller’ ( CGG); 5 §§ ‘Nicaragua (-See), Insel Ometepe, Altagracia, 11°30′n. B., 85°42′w. L., 35 m ü NN, 22. 2. 1998, leg. B. Bembé’ ( ZSM); 64 §§ ‘ COSTA RICA: Guanacaste, Palo Verde Nat’l Park, 34 km SW Canas, 1 February 1984, at eugenol, S. A. Cameron’ ( SEC); 10 §§ ‘ COSTA RICA: Guanacaste, Palo Verde Nat’l Park, 34 km SW Canas, 1 February 1984, at methyl salicylate, S. A. Cameron’ ( SEC); 7 §§ ‘ COSTA RICA: Guanacaste, Palo Verde Nat’l Park, 34 km SW Canas, 1 February 1984, at cineole, S. A. Cameron’ ( SEC); 3 §§ ‘ COSTA RICA: San José, San Antonio de Escazu , 26 March 1984, at cineole, Sydney A. Cameron’ ( SEC); 3 §§ ‘ COSTA RICA: Heredia (La Selva Biol. Res.) nr. Puerto Viejo, 3–10 Mar 1984, at cineole, Sydney A. Cameron’ ( SEC); 2 §§ ‘ COSTA RICA. San José Pro., 4 km E. San Ignacio de Acosta , 8 July 1963, 4000ft. (C. D. Michener et al.)’ ( SEC); 1 § ‘ COSTA RICA. Guanacaste Prov., El Coco, 21 June 1963 (C. D. Michener)’ ( SEC); 1 § ‘ COSTA RICA. San José Pro., San José, 13 August (Michener & Kerfoot)’ ( SEC); 35 §§ ‘ COSTA RICA: San José Prov., San Antonio de Escazú , 13–17 Apr 1985, W. T. Wcislo’ ( SEC); 7 §§ ‘ COSTA RICA: San José Prov., San Antonio de Escazú , 22–24 Apr 1985, W. T. Wcislo’ ( SEC); 7 §§ ‘ COSTA RICA: Puntarenas Prov., Monteverde, 1500 m. January 1978, visiting Ipomoea, L. A. Real’ ( SEC); 3 §§ ‘ COSTA RICA: Heredia Santo Domingo , Hotel Bougainvillea , 9°58′30″N, 84°4′36″W, 14 May 1999, D. Brzosca, CR1B99 004 ex: on vervain’ ( SEC); 1 § ‘ COSTA RICA: Alajuela ... 10°16′24″N, 84°31′24″W, 16 May 1999, D. Brzosca, CR1B99 006 attracted to methyl salicylate/ eucalyptus oil’ ( SEC); 2 §§ ‘ San José, Costa Rica’ ( ZSM); 1 § ‘ Prov. San José, Costa Rica, SN-Jeronimo- Sabanilla, 21-V-1967, col. Wille’ ( ZSM); 4 §§ ‘ San José, Costa Rica’ (SI); 1 § ‘ Finca Gibraltar, Spt. 1910, MEX.’ (SI) GoogleMaps .
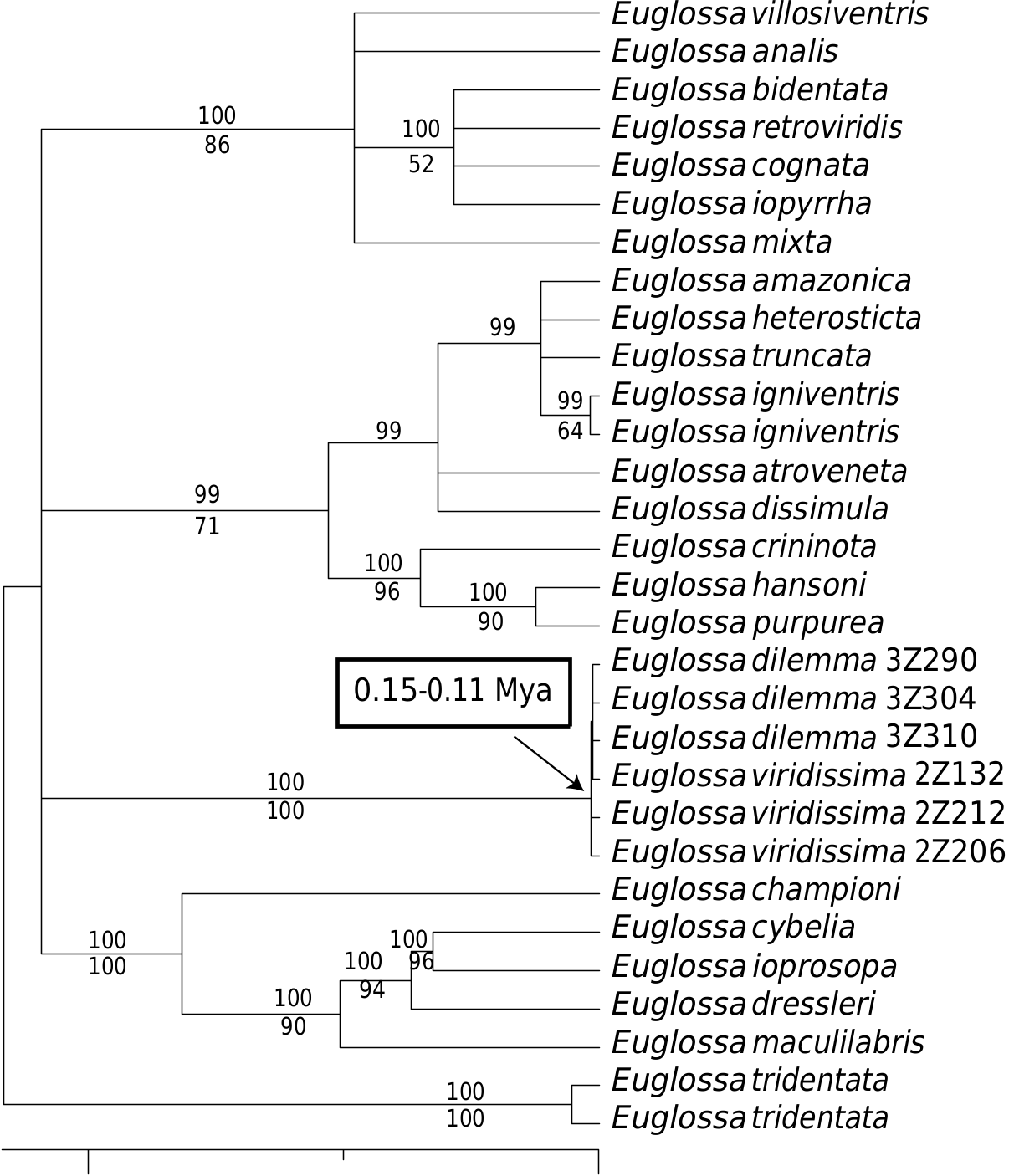
PHYLOGENETIC ANALYSIS
Both parsimony and Bayesian analyses supported the monophyly of E. viridissima + E. dilemma with high bootstrap and posterior Bayesian support ( Fig. 6 View Figure 6 ). However, neither lineage was recovered as monophyletic due to the lack of parsimony-informative characters within lineages. Only eight characters (from a total of 889) were variable between the two lineages, but all of these were parsimony-uninformative. The phylogenetic position of E. viridissima + E. dilemma within the genus Euglossa was congruent with the placement recently proposed based on several nuclear and mitochondrial genes ( Ramírez et al., 2010b). We estimated the age of the MRCA of both E. dilemma and E. viridissima by implementing a molecular clock analysis. Our results suggest that these two lineages shared a MRCA between 110 000 and 150 000 years ago. Because our phylogenetic analysis based on CO1 did not support these two lineages as reciprocally monophyletic, the MRCA age estimates represent a maximum bracket for the actual divergence of these two species.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |