
Milnesiidae Ramazzotti, 1962
|
publication ID |
https://doi.org/10.11646/zootaxa.4604.3.6 |
|
publication LSID |
lsid:zoobank.org:pub:40A4C93F-300B-46A2-B16F-DCC3B88ABF8A |
|
persistent identifier |
https://treatment.plazi.org/id/03C03821-DF13-FFD7-3AEB-8B8E1A12CE89 |
|
treatment provided by |
Plazi |
|
scientific name |
Milnesiidae Ramazzotti |
| status |
|
Family Milnesiidae Ramazzotti View in CoL , 1 962
Genus Milnesium Doyère, 1840
Milnesium cf. granulatum Ramazzotti, 1962 (elevated to species level by Michalczyk et al. 2012a)
Morphology and Morphometrics. Morphometric data for 13 adult female Milnesium cf. granulatum and six hatchlings from New Hampshire are shown in Table 2 View TABLE 2 , and for five adult females from Newfoundland in Table 3. No males were found at either site. No exuviae or eggs were found in Newfoundland. New Hampshire adults ( Figure 1A View FIGURE 1 ) have dorsal-lateral cuticle covered with a reticular design of polygons, most evident in caudal and medial regions ( Figure 2A View FIGURE 2 ). This cuticular ornamentation is sometimes difficult to see in large specimens viewed with PCM.
Pseudoplates, delineated geometric areas of cuticle ( Morek et al. 2016), are only clearly evident in the three most caudal segments ( Figure 2C View FIGURE 2 ); the pattern is the same in all individuals observed. The pattern is similar to Mil- nesium variefidum ( Morek et al. 2016) on the first segment immediately posterior to legs III (a singular rectangular paired pseudoplate flanked on each side by a smaller dorso-lateral pseudoplate), and on the second pseudoplate posterior to legs III (a cluster of eight dorsal pseudoplates flanked on each side by a dorso-lateral pseudoplate), but differs on the segment immediately above legs IV, where in Milnesium cf. granulatum the pseudoplate is not paired, but rather divided longitudinally into four parts.
Two lateral and six peribuccal lamellae are present. The buccal tube is wide ( Figure 3A View FIGURE 3 ) in full-size adults, but narrower in younger specimens, especially hatchlings ( Figure 3B View FIGURE 3 ); i.e., buccal tube width is significantly allometric ( Table 2 View TABLE 2 ). In Newfoundland significant allometry was not detected in buccal width ( Table 3). Fewer specimens, representing a narrower range of body lengths, were available in the Newfoundland samples, making detection of allometry less likely. The adult claw formula is [3-3]-[3-3] ( Figure 4A, C View FIGURE 4 ).
Thirty out of 38 eggs hatched within 10 days of being placed in water ( Figure 1B View FIGURE 1 ). Hatchlings had the same reticular cuticular ornamentation as adults ( Figure 2B View FIGURE 2 ). Claw configuration in hatchlings was [3-3]-[3-3] ( Figure 4B, D View FIGURE 4 ), identical to their mothers ( Figure 4A, C View FIGURE 4 ).
Genetic analysis. The COI gene was amplified from 11 New Hampshire specimens and 7 Newfoundland specimens. New Hampshire adults exhibited a genetic diversity of 6 haplotypes with p-distances of 0.2–0.7% over a 581 bp region while Newfoundland adults displayed a genetic diversity of 3 haplotypes with p-distances of 0.2–0.3% over a 604 bp region of the COI gene (see Table 1 View TABLE 1 for accession numbers). Milnesium cf. granulatum from New Hampshire and Newfoundland exhibited a total of 6 COI haplotypes and the two populations had a p-distance of 0.2% further suggesting that they represented the same species.
Genetic distance of the COI gene was compared between Milnesium cf. granulatum and other Milnesium spp. and were as follows: Milnesium berladnicorum ( Ciobanu et al. 2014; from Romania)—18%, Milnesium tardi- gradum (Michalczyk et al. 2012a; from Germany)—18%, Milnesium variefidum ( Morek et al. 2016; from Scotland)—18%, and Milnesium lagniappe (Meyer et al. 2013; from Louisiana, USA)—21%. The overall mean distance in the COI gene for Milnesium spp. was 17%. Among Milnesium spp., Milnesium berladnicorum and Milnesium variefidum had the lowest p-distance (13%) while several species had p-distances of 21% ( Table 4 View TABLE 4 ).
)
A 564 bp fragment of the ITS-2 region was also amplified from 20 adult Milnesium cf. granulatum specimens from New Hampshire. Molecular analysis was performed on a 407 bp region of the fragment using MEGA7. Tajima’s Neutrality Test indicated population expansion occurring in the animals collected for analysis. The number of identified ITS-2 haplotypes was 13, with p-distances ranging from 0.2–3.7% with an overall mean distance of 1.6% (see Table 1 View TABLE 1 for accession numbers). The ITS-2 region of siblings from 8 clutches was also analyzed. Each clutch contained between 2 and 5 siblings. Genetic data demonstrated that 2–5 haplotypes were present in siblings from a single clutch ( Table 5 View TABLE 5 ). DNA sequencing for most hatchlings resulted in shorter DNA sequences varying in length between 111 bp and 440 bp (269 bp on average) allowing for fewer bases to be analyzed. Genetic distance of ITS- 2 sequences was compared between Milnesium cf. granulatum and Milnesium spp. available in GenBank ( Table 6 View TABLE 6 ). The p-distances were as follows: Milnesium berladnicorum —22 %, Milnesium tardigradum —20%, Milnesium variefidum —21%, and Milnesium lagniappe —27%. The overall mean distance in the ITS-2 region for Milnesium spp. was 26%. Among Milnesium spp., Milnesium berladnicorum and Milnesium variefidum had the lowest p-distance (9%) while Milnesium lagniappe and Milnesium tardigradum had the highest p-distance (28%). A phyloge- netic tree of Milnesium spp. and two Echiniscus spp. was constructed using the neighbor-joining method. Bootstrap values were high for all branches (95–100%) except one (50%), which was located at the tip of the phylogenetic tree ( Figure 5 View FIGURE 5 ).
[1] [2] [3] [4]
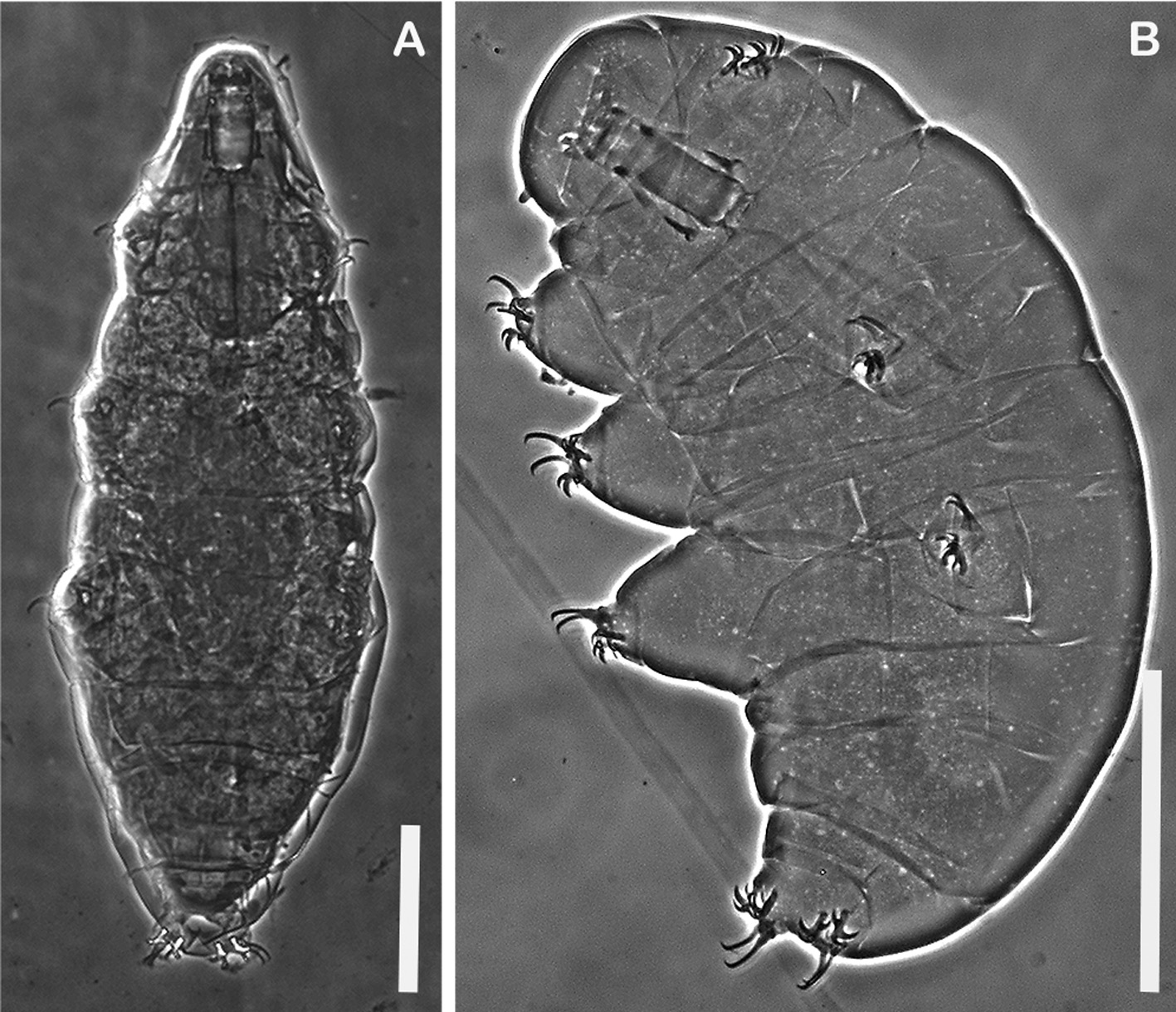
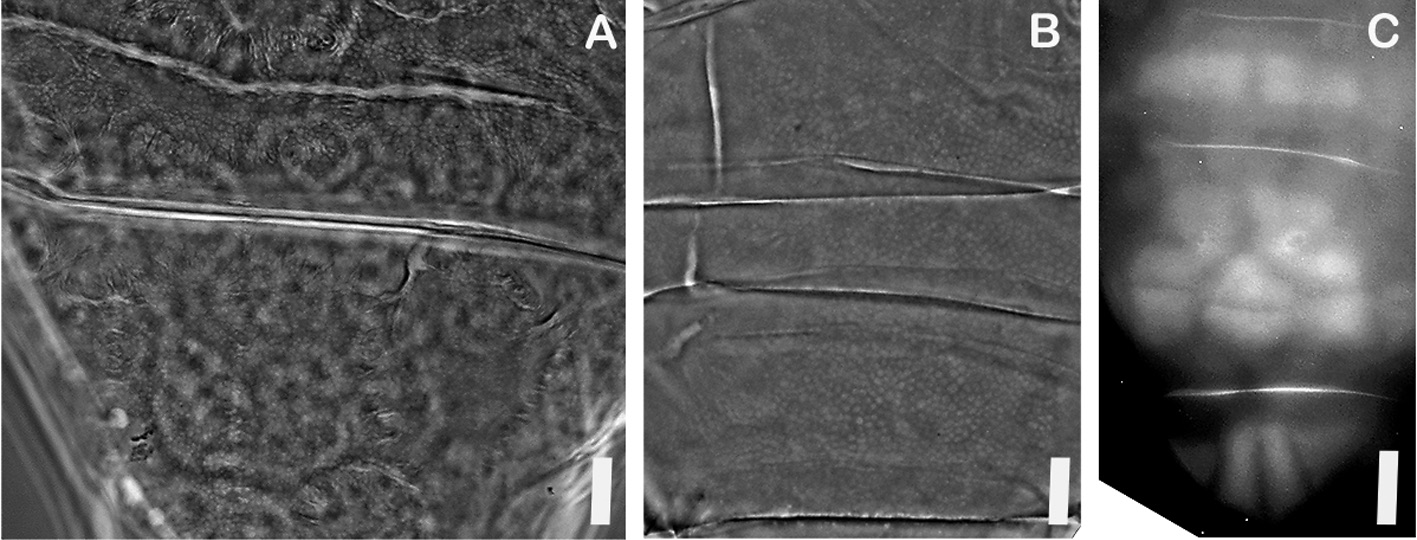

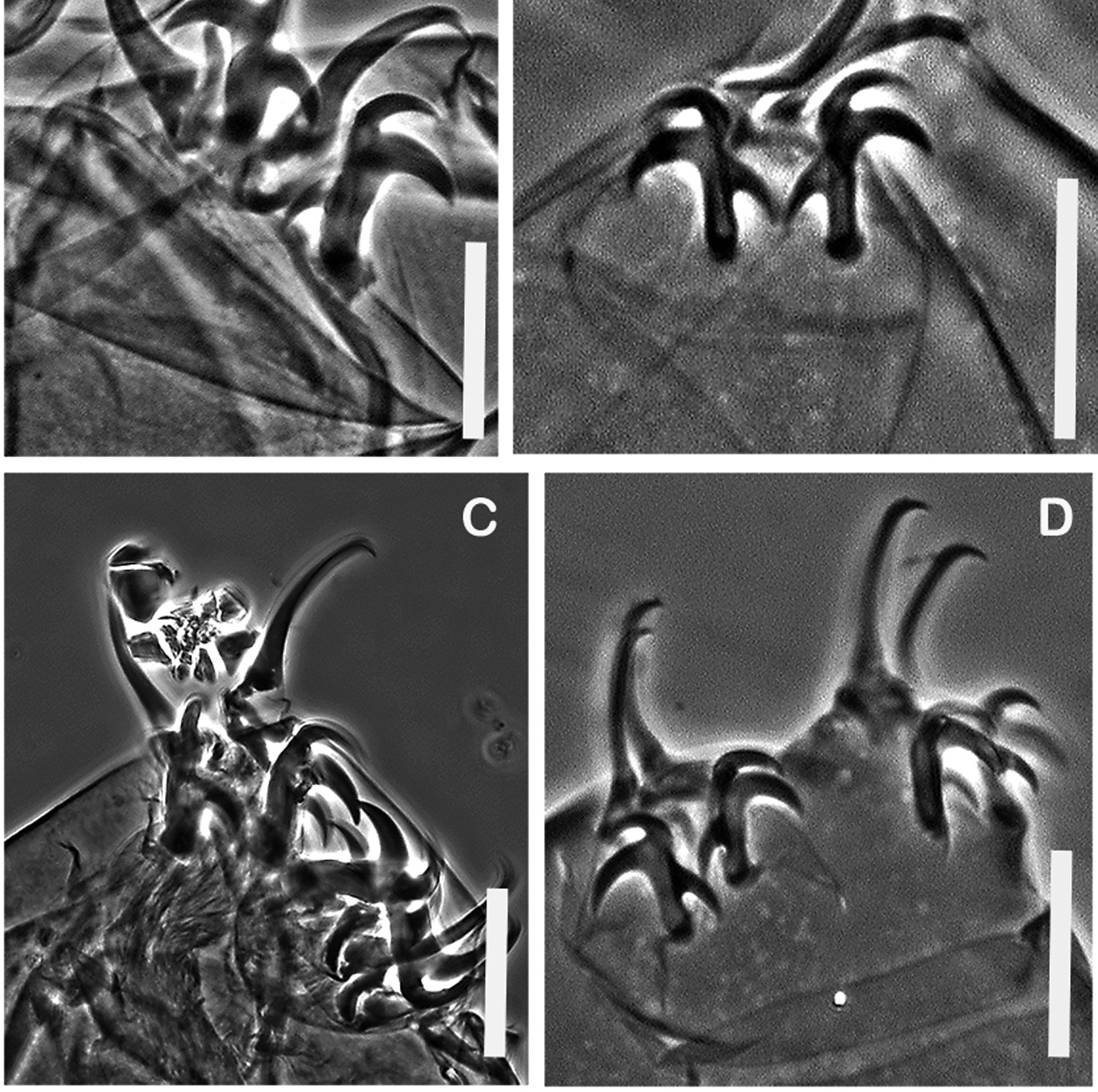
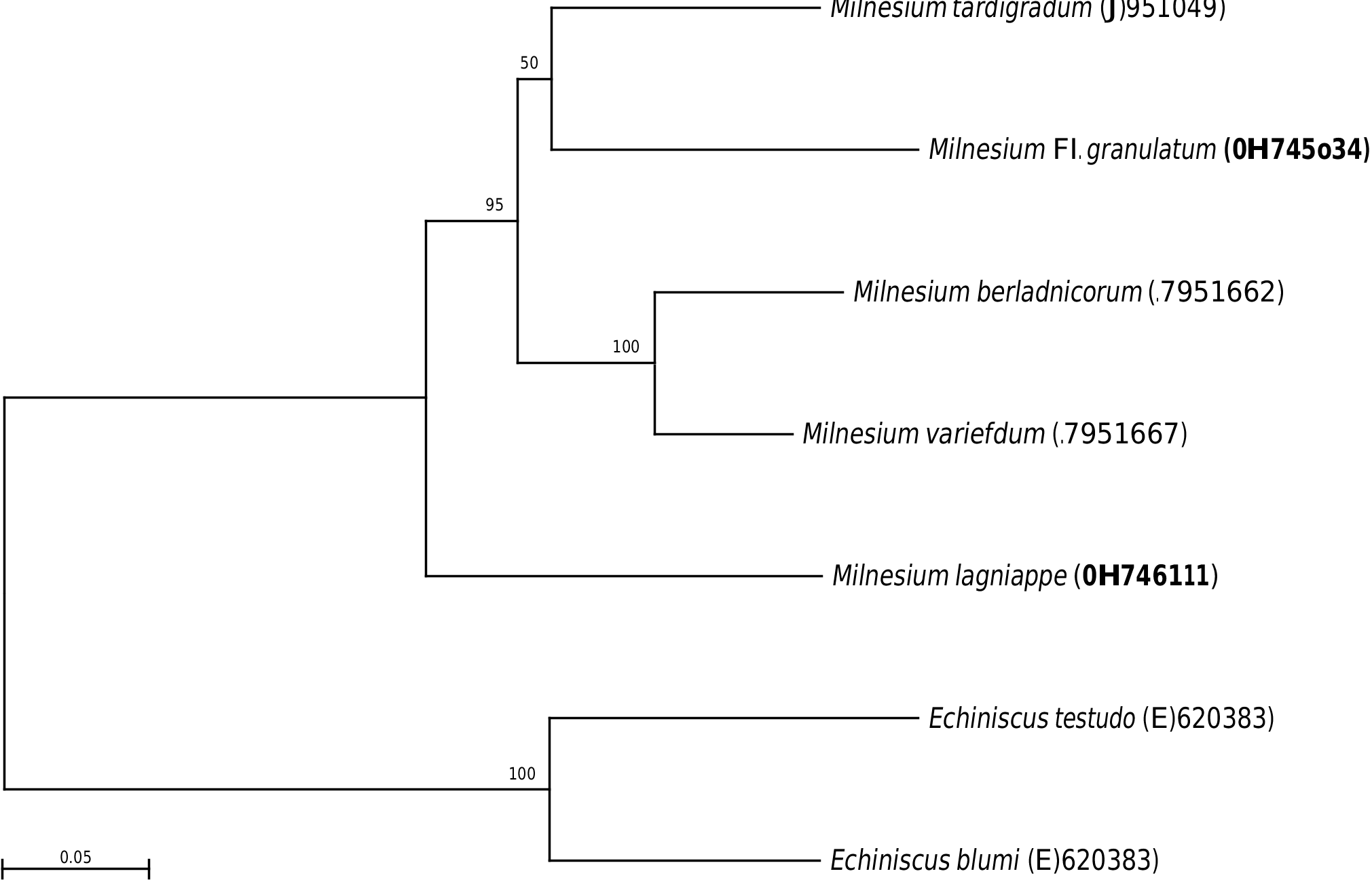
TABLE 2. Morphometric data and pt values of selected characters of 13 adult females and six hatchlings of Milnesium cf. granulatum from New Hampshire, USA. Range refers to the smallest and largest structure found among all measured specimens. All lengths in micrometers. Abbreviations: N = number of specimens measured, SD = standard deviation, pt = pt index, b = allometric exponent and a* = Y-intercept of the regression line of the Thorpe normalized trait versus buccal tube length. Values of b followed by an asterisk indicate that the character was significantly allometric (P <0.05). a* pt was calculated using the mean value of buccal tube length.
| Character | N | Range | Mean | SD | b | a* | a*pt | |||
|---|---|---|---|---|---|---|---|---|---|---|
| μm | pt | μm | pt | μm | pt | μm | ||||
| Body length | 19 | 245–773 | 819 –1486 | 455 | 1161 | 188 | 213 | 1.32* | 453 | 1185 |
| Peribuccal papilla length | 13 | 4.5–11.9 | 15.5–28.6 | 8.2 | 21.2 | 2.8 | 3.8 | 1.06 | 8.4 | 22.0 |
| Lateral papilla length | 10 | 4.5–12.4 | 11.7–23.7 | 7.4 | 17.8 | 2.4 | 3.3 | 0.82 | 6.6 | 17.3 |
| Buccal tube | ||||||||||
| Length | 19 | 27.7–54.2 | – | 38.2 | – | 11.2 | – | – | – | – |
| Stylet support insertion point | 18 | 17.8–34.5 | 61.9–65.2 | 24.7 | 63.7 | 7.0 | 1.0 | 0.97* | 24.3 | 63.6 |
| Anterior width | 19 | 13.5–30.1 | 45.6–55.7 | 19.7 | 51.1 | 6.5 | 3.0 | 1.11* | 19.6 | 51.3 |
| Standard width | 18 | 11.9–28.3 | 40.2–52.2 | 18.6 | 47.0 | 6.6 | 3.7 | 1.23* | 18.0 | 47.1 |
| Posterior width | 17 | 11.6–21.9 | 37.7–43.4 | 15.6 | 41.3 | 6.6 | 3.7 | 0.92* | 15.7 | 41.1 |
| Standard width/length ratio | 18 | 40%–52% | – | 47%. | – | 4% | – | – | 47% | – |
| Posterior/anterior width ratio | 17 | 68%–93% | – | 81% | – | 6% | – | – | 80% | – |
| Claw 1 lengths | ||||||||||
| External primary branch | 18 | 8.4–16.0 | 28.5–40.7 | 12.2 | 33.2 | 2.8 | 3.7 | 0.76* | 12.6 | 33.0 |
| External base + secondary branch | 19 | 7.6–14.3 | 22.3–32.5 | 10.4 | 27.5 | 2.6 | 2.3 | 0.82* | 10.4 | 27.2 |
| External spur | 19 | 2.7–7.0 | 9.4–15.2 | 4.4 | 11.6 | 1.2 | 1.5 | 0.84 | 4.4 | 11.5 |
| External branches length ratio | 18 | 70%–97% | – | 83% | – | 7% | – | – | 83% | – |
| Internal primary branch | 19 | 8.9–16.0 | 27.9–37.9 | 12.1 | 32.4 | 2.7 | 2.8 | 0.75* | 12.2 | 31.9 |
| Internal base + secondary branch | 19 | 7.8–14.6 | 22.9–33.9 | 10.5 | 27.9 | 2.6 | 2.4 | 0.81* | 10.6 | 27.7 |
| Internal spur | 18 | 2.8–8.0 | 9.7–16.2 | 4.9 | 12.4 | 1.7 | 1.6 | 1.12 | 4.8 | 12.6 |
| Internal branches length ratio | 19 | 74%–96% | – | 86% | – | 6% | – | – | 87% | – |
| Claw 2 lengths | ||||||||||
| External primary branch | 18 | 9.2–17.9 | 30.0–40.8 | 13.1 | 34.7 | 3.0 | 3.6 | 0.75* | 13.1 | 34.3 |
| External base + secondary branch | 15 | 7.6–14.5 | 24.4–32.7 | 10.7 | 27.8 | 2.5 | 2.7 | 0.78* | 10.6 | 27.7 |
| External spur | 12 | 3.3–6.6 | 11.1–14.3 | 5.1 | 12.4 | 1.3 | 1.0 | 0.91 | 4.8 | 12.6 |
| External branches length ratio | 15 | 73%–88% | – | 81% | – | 5% | – | – | 81% | – |
| Internal primary branch | 18 | 9.4–20.2 | 30.4–39.9 | 13.2 | 34.6 | 3.5 | 2.7 | 0.85* | 13.1 | 34.3 |
| Internal base + secondary branch | 17 | 7.7–14.4 | 21.0–32.8 | 10.9 | 28.0 | 2.6 | 2.8 | 0.77* | 10.6 | 27.7 |
| Internal spur | 16 | 2.8–7.9 | 10.0–15.7 | 5.3 | 13.3 | 1.7 | 1.7 | 1.09 | 5.2 | 13.6 |
| Internal branches length ratio | 16 | 69%–90% | – | 80% | 6% | – | – | 81% | – | |
......continued on the next page
TABLE 4. P-distances in the COI gene between Milnesium cf. granulatum and several Milnesium species.
| [1] | [2] | [3] | [4] | |
|---|---|---|---|---|
| [1] Milnesium cf. granulatum | ||||
| [2] Milnesium lagniappe | 20.5% | |||
| [3] Milnesium berladnicorum | 17.7% | 20.7% | ||
| [4] Milnesium variefidum | 18.1% | 18.9% | 12.6% | |
| [5] Milnesium tardigradum | 18.1% | 20.5% | 14.3% | 13.8% |
TABLE 5. Genetic diversity of Milnesium cf. granulatum siblings from 8 clutches expressed as the number of identified ITS-2 haplotypes.
| Clutch | Number of individuals per clutch | Number of haplotypes | Sequence read length |
|---|---|---|---|
| 1 | 3 | 3 | 111 bp |
| 2 | 5 | 4 | 280 bp |
| 3 | 2 | 2 | 440 bp |
| 4 | 5 | 4 | 176 bp |
| 5 | 4 | 3 | 314 bp |
| 6 | 2 | 1 | 259 bp |
| 7 | 3 | 2 | 185 bp |
| 8 | 4 | 4 | 387 bp |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Milnesiidae Ramazzotti
| Jackson, Kathy Jo A. & Meyer, Harry A. 2019 |
Milnesium cf. granulatum
| Ramazzotti 1962 |