
Longior similis Morffe, García & Ventosa, 2009
|
publication ID |
https://doi.org/10.11646/zootaxa.4399.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:879B3B11-4716-4F64-8177-8B11515C23F0 |
|
DOI |
https://doi.org/10.5281/zenodo.5946629 |
|
persistent identifier |
https://treatment.plazi.org/id/03C1879D-FFA6-FFCC-A8AC-FCA62850F832 |
|
treatment provided by |
Plazi |
|
scientific name |
Longior similis Morffe, García & Ventosa, 2009 |
| status |
|
Longior similis Morffe, García & Ventosa, 2009
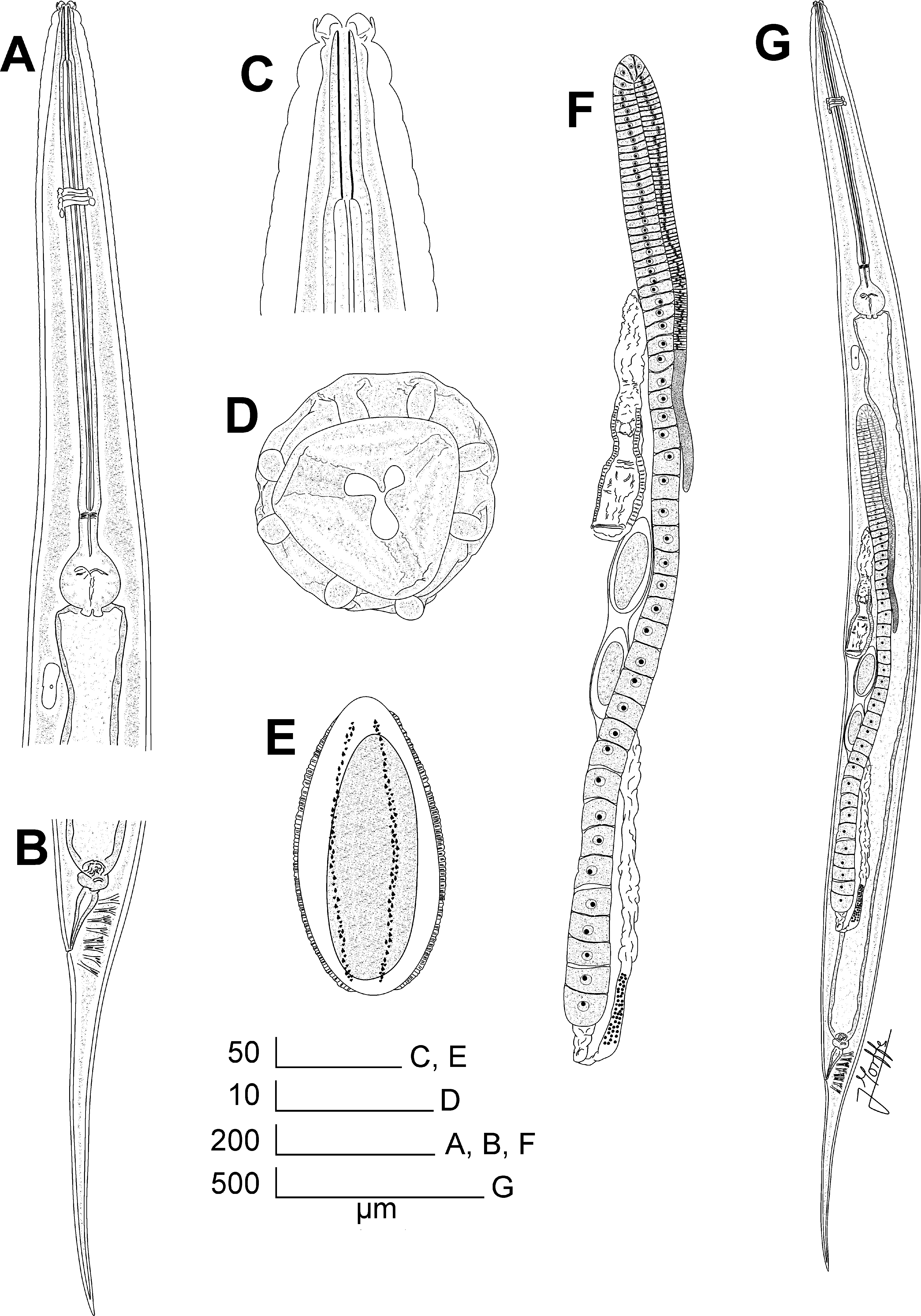
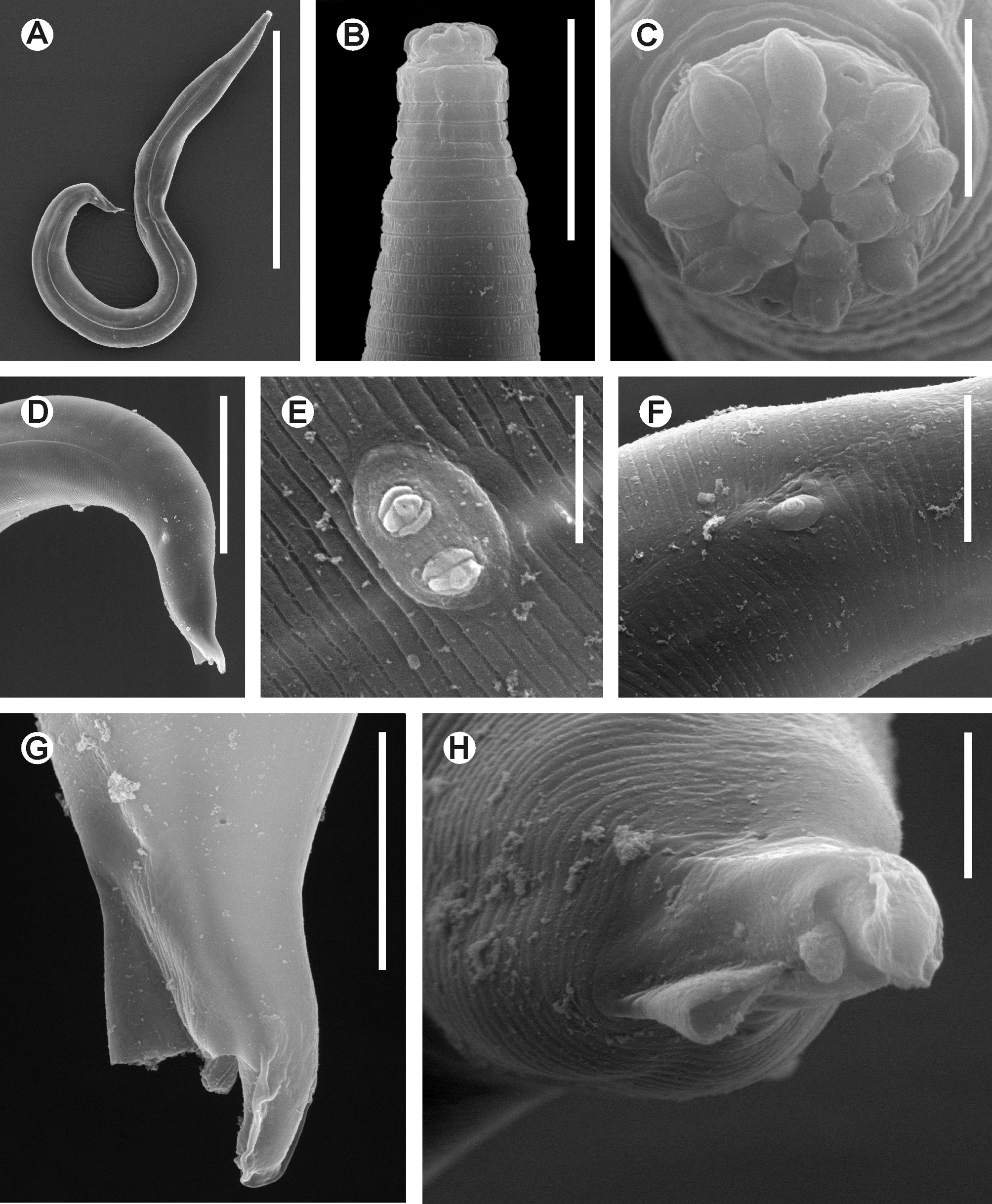
Fig. 5 A–G View FIGURE 5 , Fig. 6 A–G View FIGURE 6 , Fig. 7 A–F View FIGURE 7 , Fig. 8 A–H View FIGURE 8
Longior similis Morffe, García & Ventosa, 2009: 14 –16, fig. 1. A–G
Material examined. Vouchers : 9♀♀, Cuba, Artemisa province, Candelaria, Sierra del Rosario, El Salón; in Passalus interstitialis ; 12/VII/2014; J. Morffe, N. García coll.; CZACC 114956–11.4964 . 9♂♂, same data as the latter; CZACC 11.4965–11.4973 . 2♀♀, same data as the latter; MNHNSD 05.003–05.004 . 2♂♂, same data as the latter; MNHNSD 05.005–05.006 . 2♀♀, same data as the latter; CHIOC 38416 View Materials a – b. 2♂♂, same data as the latter; CHIOC 38416 View Materials c–d.
Vouchers: 17♀♀, Cuba, Isla de la Juventud, Sierra de Casas; in Passalus interstitialis ; IV/2014; J. Morffe, N. García, M. Olcha coll.; CZACC 11.4989–11.4998, 11.5000–11.5006. 11♂♂, same data as the latter; CZACC 11.5007–11.5017.
Vouchers: 2♀♀, Cuba, Sancti Spíritus province, Trinidad, Topes de Collantes , Hotel “ Los Helechos ”; in Passalus interstitialis ; V/2013; E. Fonseca coll.; CZACC 11.4730–11.4731 .
Vouchers: 8♀♀, Cuba, Sancti Spíritus province , Trinidad, Topes de Collantes, path to the River Caburní ; in Passalus interstitialis ; 12/X/2014; J. Morffe, N. García coll.; CZACC 11.4925–11.4932 . 2♂♂, same data as the latter; CZACC 11.4933–11.4934 .
Vouchers: 4♀♀, Cuba, Ciego de Ávila province, Chambas, Sierra de Jatibonico , La Piedra ; in Passalus interstitialis ; 25/XI/2014; C. Martínez, R. Cardet coll.; CZACC 11.4891–11.4894 . 3♂♂, same data as the latter; CZACC 11.4895–11.4897 .
Redescription. Female. Body long and slender, widening gradually posterior to head, reaching its maximum width at level of the vulva, then narrowing gradually towards tail. Cuticle thin. Sub-cuticular striae present. Cervical cuticle unarmed, with wide annuli ( ca. 8 µm) from the base of the first cephalic annule to ca. the level of the first third of procorpus. Lateral alae well developed, extend from ca. two body-widths posterior to the basal bulb to the level of the anus. Head bearing eight paired, digitiform cephalic papillae, which originate from the external edge of the head, at ca. half of its height. A cuticular, triradiate veliform annular lip surrounds the oral opening. Amphids lateral, at level of the base of the cephalic papillae. First cephalic annule comparatively long, truncate, slightly dilated, ca. 1.5–2 head-lengths long. Stoma long and slender, surrounded by an oesophageal collar. Oesophagus consists of a muscular, sub-cylindrical procorpus, its base similar in diameter to the isthmus. Basal bulb pyriform, valve-plate well developed. Intestine simple, sub-rectilinear, its fore region slightly dilated. Rectum comparatively long. Anus as a crescent-like ventromedian slit, anteriorly directed, not prominent. Nerve ring encircling procorpus at ca. 30% of its length. Excretory pore located at ca. two thirds to a body-width posterior to the basal bulb. Vulva a median transverse slit ca. 45 µm in length, its lips not prominent, located at level of midbody. Anterior lip of vulva slightly overlapping the posterior. Vagina muscular, forwardly directed. Genital tract monodelphic-prodelphic, occupying ca. 40% of the body length. Ovary distally reflexed at ca. a body-width posterior the excretory pore, distal flexure ca. 1.5 body-widths in length. Oocytes in a single row. Eggs ellipsoidal, bearing eight rough longitudinal ridges in the shell that do not reach the poles. A number of eggs, 1–9 at a time in the uterus (more frequently 1– 3 eggs). Phasmids pore-like, lateral, barely evident, located ca. 8 µm posterior to the end of lateral alae. Tail conical, subulate, ending in a fine tip.
Male. Body comparatively shorter and more slender than that of females. Posterior region ventrally curved in heat-fixed specimens. Cuticle unarmed, finely annulated, from the base of the head to the level of the sub-lateral pair of copulatory papillae. Annuli more evident and wide ( ca. 2 µm) at level of the cervical region, until the beginning of lateral alae. Annuli at level of the sub-lateral pair of copulatory papillae ca. 1 µm in width. Lateral alae narrow, extend from the oesophageal region, at level of the procorpus ( ca. its midpoint) to ca. the midpoint between the median and sub-lateral pairs of copulatory papillae. Head set-off from body by a deep groove, bearing eight ellipsoidal, slightly flattened cephalic papillae; arranged as two sub-dorsal and two sub-ventral pairs. Six digitiform lips originating from the internal edge of the cephalic papillae and projecting to the center of the oral aperture. Lips arranged as one lip dorsal, one ventral, two sub-dorsal and two sub-ventral. Sub-dorsal and subventral lips flank the dorsal and ventral lips, respectively. Amphids lateral, as crescent-like pores. Oesophagus consists of a muscular, sub-cylindrical procorpus, diminishing its diameter toward its posterior half, almost equaling the diameter of the isthmus at level of their junction. Basal bulb rounded, valve-plate well developed. Intestine simple, its fore region slightly inflated. Nerve ring encircling procorpus at ca. 45% of its length. Excretory pore located at ca. a body-width posterior to the basal bulb. Monorchic, testis outstretched. Vas deferens with three distinguishable regions: an anterior region with granular content; a median, slightly swollen region, also granular (granules slightly shorter in diameter) and a posterior region that diminishes its diameter towards the cloaca. Spicule absent. A ventral bursa-like structure present. Dorsal cuticle of the posterior end thickened and smooth from the tail tip to ca. the midpoint between the ventromedian pair of papillae and tail tip. Such thickening forms a cuticular crest at the tail tip that extends terminally and laterally until the level of the distal end of bursa. Phasmids pore-like, lateral, located at level of the beginning of the bursa. Four pairs of copulatory papillae, two pre-cloacal and two post-cloacal. Ventromedian pre-cloacal larger pair consisting of duplex papillae very close to each other on an ellipsoidal protuberance (appear to be a single papilla in lateral view) at ca. 80 µm from the tail tip. One sublateral pre-cloacal pair of papillae are present at ca. the midpoint between the ventromedian pair and the tail tip. Remaining two post-cloacal pairs consist of small papillae: one pair ventral, located at the tip of a papilliform protuberance just posterior to the bursa and one pair ventral, sub-terminal, close to the tail tip, below and in contact with the terminal cuticular crest. Posterior end slightly curved dorsally at level of the bursa-like structure.
Remarks. The description of the male of L. similis was based on a single specimen ( Morffe & García 2011) and the differences with L. longior were based only on the presence of annuli on the cervical cuticle and the length of the oesophagus. As mentioned above, it is inadvisable to consider the annuli of the cervical region as diagnostic. On the other hand, a larger number of specimens were examined for the present study, obtaining a more representative sample for morphometric comparisons. Analysis of these measurements reveal that males of L. similis have a shorter body than L. longior (total length = 0.860–1.210 vs. 1.075–1.590, measurements of this study). The oesophagus of L. similis is, indeed, slightly shorter than L. longior (b = 3.37–4.00 vs. 2.97–3.77).
The lateral alae were not mentioned in the original description, nor considered in the differential diagnosis, but differ in their extension. In L. similis the lateral alae extend from ca. the midpoint of the procorpus to ca. the midpoint between the median and sub-lateral pairs of copulatory papillae vs. extending from the level of the last third of the procorpus to ca. half of the body width before the level of the median pair of copulatory papillae. Both taxa present a similar arrangement of the copulatory papillae and the main differences are in the shape of the tail end: slightly curved dorsally in L. similis and straight in L. longior .
El Salón, Sierra del Rosario constitutes the first locality record of L. similis in Artemisa province in Western Cuba. On the other hand, the discovery of the species in Hotel “Los Helechos” and Caburní river, in the Gran Parque Natural Topes de Collantes and Las Piedras, Sierra de Jatibonico; extends its distribution to Sancti Spíritus and Ciego de Ávila provinces respectively, being also the first records for Central Cuba. Out of the main island of the archipelago, L. similis was recorded for Sierra de Casas in the northern part of Isla de la Juventud. Previously, L. similis was only known from Escaleras de Jaruco (its type locality), Mayabeque province and El Pan de Matanzas, Matanzas province, both localities located in Western Cuba ( Morffe & García 2010; Morffe et al. 2009). The individuals from the aforementioned new localities coincide morphologically and morphometrically ( Tables 3 and 4) with the previous records of the species.
DNA studies. Nine partial sequences (773–816 bp) of the D2-D3 region of the 28S LSU rDNA were obtained from both L. longior (five sequences, four females and one male) and L. similis (four sequences, three females and one male). L. longior and L. similis differ at 13 homologous positions in a 641 bp sequence alignment. This supports the morphological and morphometric differences and confirms their status as different species. No intraspecific differences were recorded between the studied populations of L. longior and L. similis .
The ML and BI trees showed identical topology, thus only the ML tree is shown ( Fig. 9 View FIGURE 9 ). Both L. longior and L. similis cluster with Hystrignathus sp., the only available hystrignathid sequence, and form a well-supported clade. The three Travassosinema ( type genus of Travassosinematidae ) included in the present study and Cameronia multiovata form a clade as sister group to the hystrignathids. This is concordant with Spiridonov & Guzeeva (2009) who analyzed the phylogeny of Thelastomatoidea based on the same molecular marker. These authors also obtained a clade formed by Hystrignathus sp., Cameronia sp. and Travassosinema sp.
The identification of males among Thelastomatoidea by means of morphology and morphometrics is complex, since they are usually quite different from their conspecific females. This task becomes more difficult in cases of multi-specific infestations, where several male morphotypes can be found in a host. Several authors ( Jex et al. 2006; Spiridonov & Cribb 2012; Spiridonov & Guzeeva 2009) stated the usefulness of molecular markers such as the 28S LSU rDNA for the proper identification of thelastomatoid males. The sequences of male specimens included in the present study coincide completely with females, confirming the conspecificity of both sexes among L. longior and L. similis .
The males of L. longior and L. similis resemble those of L. longicollis , the type species of the genus, by having the oesophagus with the procorpus sub-cylindrical, similar in diameter to the isthmus. The posterior end presents a bursa-like structure and the thickened dorsal cuticle characteristic of the genus. This supports the proper identification of the males of Longior by Travassos & Kloss (1958). Nevertheless, new collections of L. longicollis are needed for SEM and molecular studies in order to obtain more accurate information on its morphology and phylogeny.



| CZACC |
Coleccion Zoologia, Academia de Ciencias de Cuba |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Thelastomatoidea |
|
Family |
|
|
Genus |
Longior similis Morffe, García & Ventosa, 2009
| Morffe, Jans, García, Nayla, Breugelmans, Karin & Adams, Byron J. 2018 |
Longior similis Morffe, García & Ventosa, 2009: 14
| Morffe, Garcia & Ventosa 2009: 14 |