
Samarplax principe, Husana, Daniel Edison M., Tan, Swee Hee & Kase, Tomoki, 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.279281 |
|
DOI |
https://doi.org/10.5281/zenodo.6182670 |
|
persistent identifier |
https://treatment.plazi.org/id/03C2082F-FF81-FF8E-FF75-FB9344A4FCA9 |
|
treatment provided by |
Plazi |
|
scientific name |
Samarplax principe |
| status |
sp. nov. |
Samarplax principe View in CoL new species
( Figs. 3–8 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 )
Material examined. Holotype. Male, 3.52 x 3.69 mm (NMCR-39033), Principe Cave, Guiuan, Eastern Samar, Philippines, 10˚58.637´N, 125˚48.373´E coll. E. Husana, 2 Nov. 2006.
Paratypes. 3 males [3.64 x 3.63 mm ( ZRC 2011.0880; ex. NSMT-Cr 2007-0003); 4.09 x 3.98 mm, used for SEM (NSMT-Cr 2007-00040); 3.22 x 3.25mm (NSMT-Cr 2007-0005)] same data as holotype. 4 females [5.35 x 5.68 mm, ovigerous ( ZRC 2011.0879; ex. NSMT-Cr 2007-0002); 4.81 x 4.78 mm, ovigerous (NSMT-Cr 2007- 006); 4.08 x 4.06 mm, immature (NSMT-Cr 2007-0007); 4.72 x 4.69 mm, used for SEM (NSMT-Cr 2007-0008)], same data as holotype.
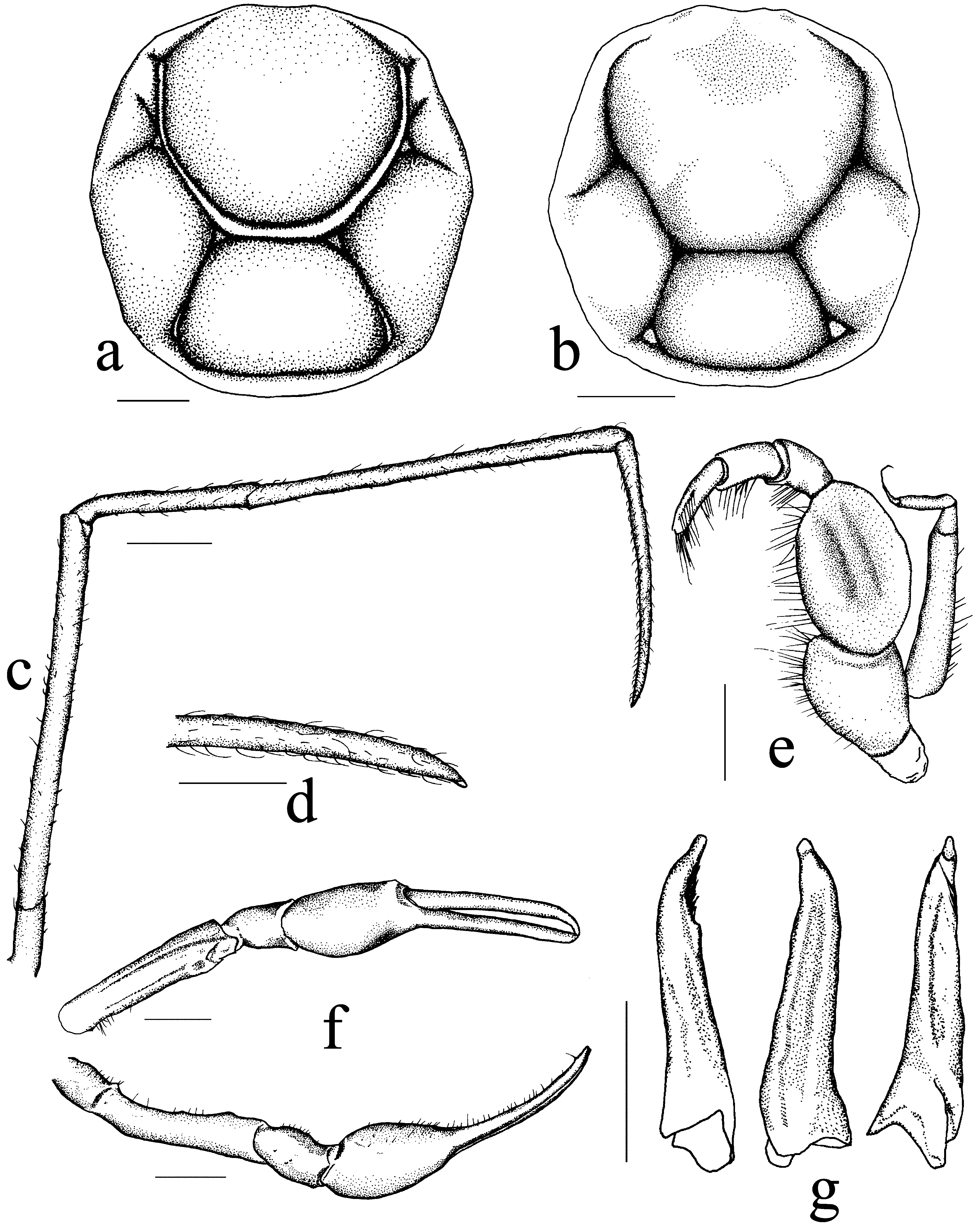
Description. Carapace sub-polygonal, about same length or longer than wide; surrounded by continuous rim. Gastrocardiac, thoracic and cervical grooves deep, distinct; thoracic grooves diverging and converging posteriorly but not reaching posterior border; cervical groove long converging anteriorly towards frontal region. Gastric, cardiac and epibranchial regions inflated, epibranchial slightly lower than gastric and cardiac regions. Rostrum absent; frontal, anterolateral, midlateral, posterolateral and posterior edges clearly demarcated. Frontal margin weakly bilobed, 2 longitudinal fissures present just below frontal margin parallel to edge; anterolateral, midlateral and posterior margins cristate; anterolateral margin entire; posterolateral margin entire. Epistome relatively broad. Branchiostegal region swollen laterally.
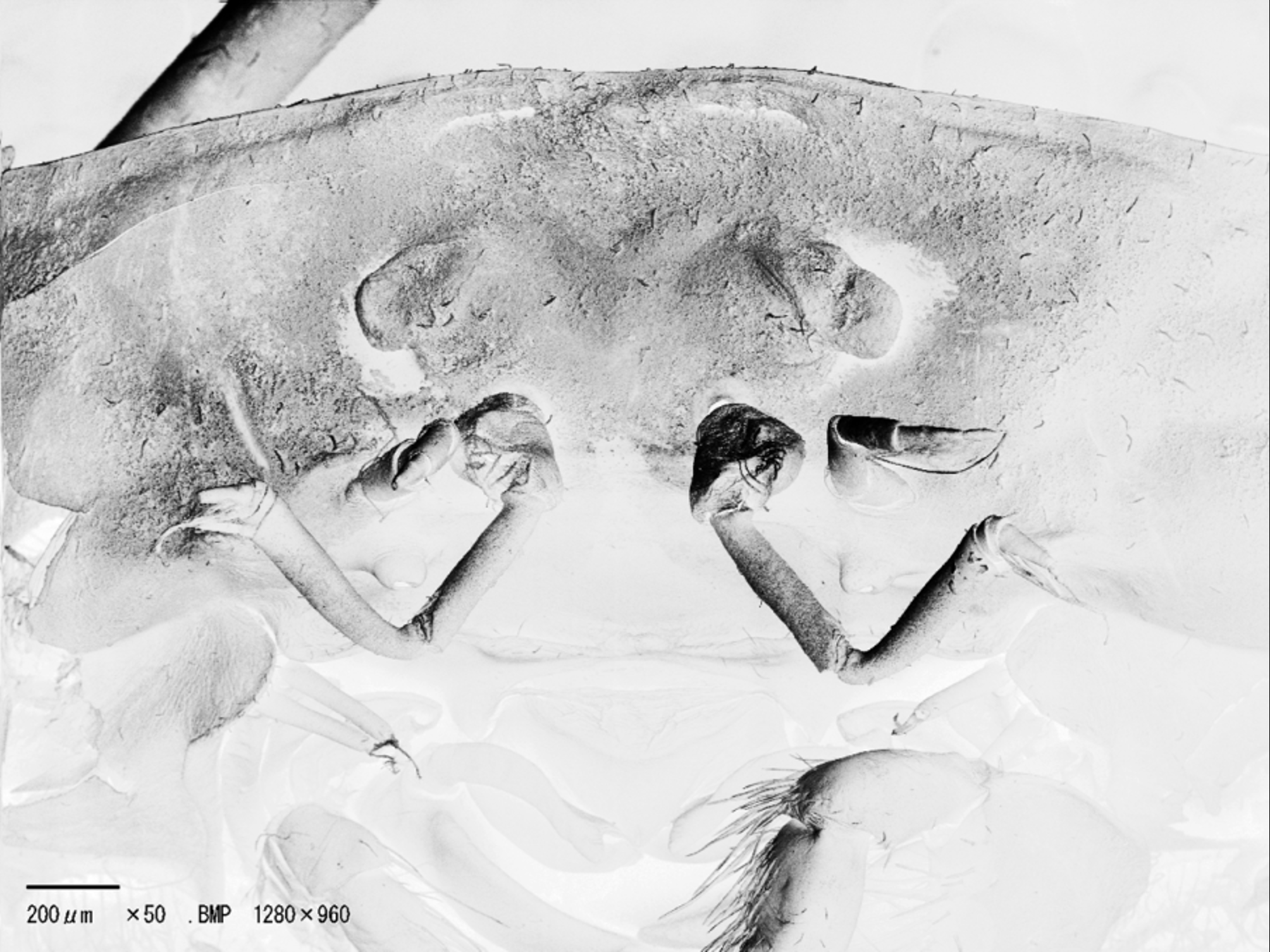
Eyes not visible dorsally; eyestalk very short, immovable, cornea absent, orbits incomplete. Antennular fossa very shallow, medially separated by thin, very low septum, basal article swollen, protruding from fossa. Antennal fossae absent, antenna well developed, basal article relatively narrow, not positioned directly below base of eye stalk, offset to outer edge.
Third maxillipeds narrow, with wide gape when closed; ischium with broad distal inner lobe; merus inner and outer margins divergent distally; merus twice length of ischium along lateral margin; medial margins of merus and ischium lined with strong setae; palp long; exopod short, reaching about two-thirds of merus, with distinct flagellum.
Chelipeds equal, slender, long; surfaces smooth; inner margins of merus lined with simple setae and submedian dorsal tooth. Carpus smooth, short, lined with setae along inner edge. Palm and finger equal in length relatively long, without teeth, lined with short setae along cutting edges.
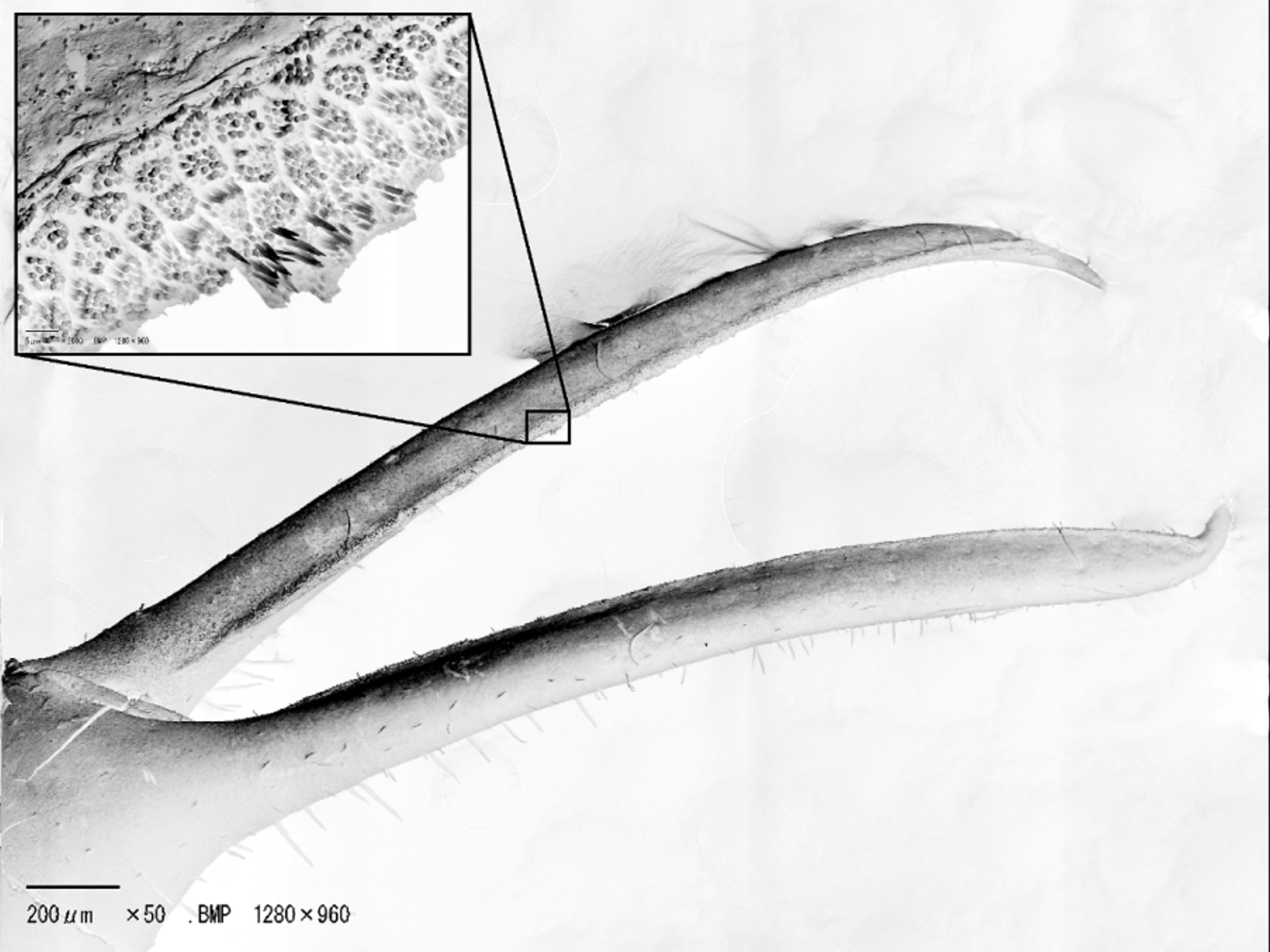
Ambulatory legs slender, long, with short scattered setae; P3 length about 4 times carapace width, slightly longer than P2, P5 not clearly shorter than rest; P2 to P5 meri slightly longer to longer than carapace length, cross-section elliptical, diameter constant throughout length; propodi shorter than respective meri; dactyli shorter than respective propodi, curved inwards slightly, margins entire.
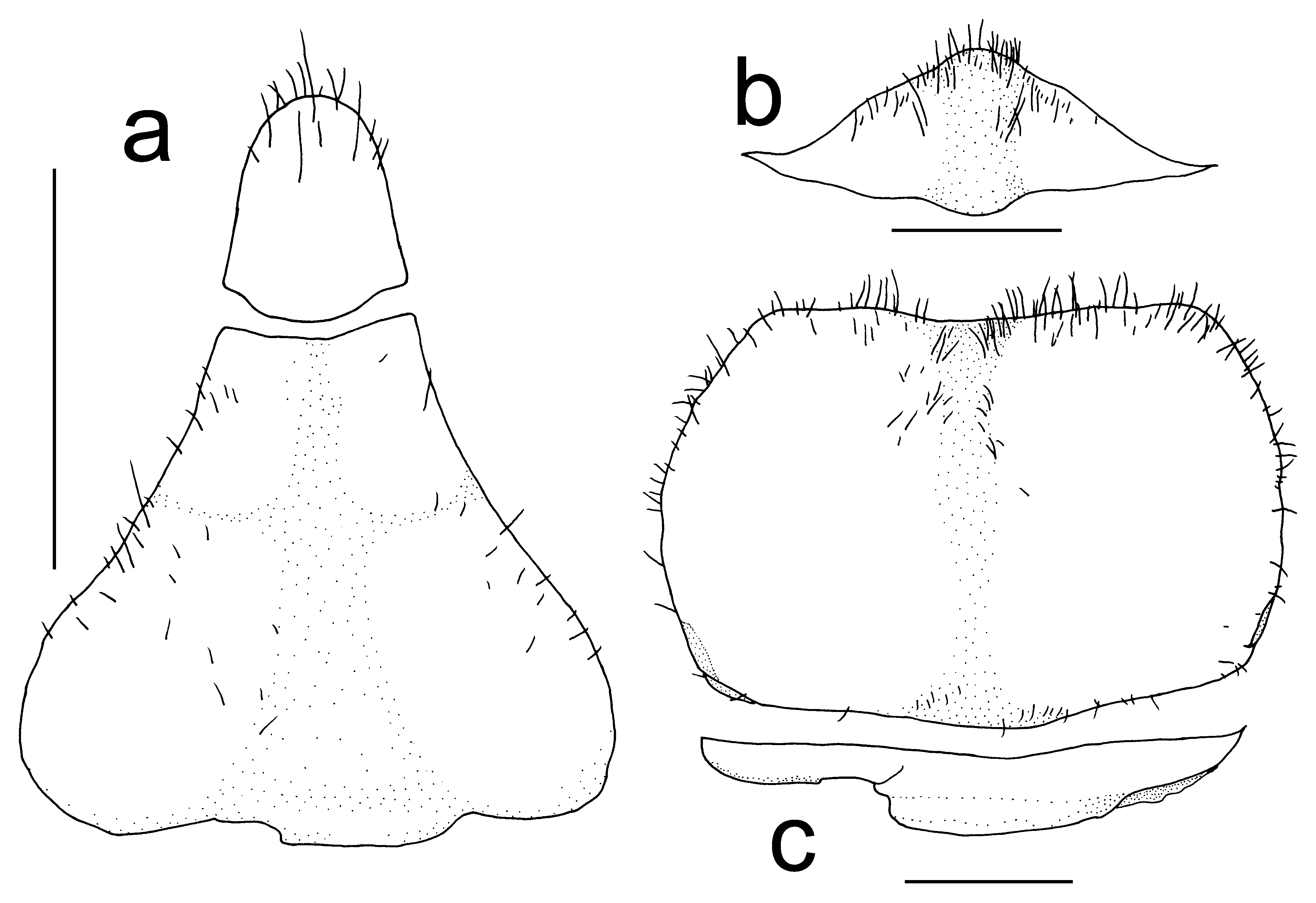
Male abdomen with 4 parts; somite 1 with proximal part below carapace posterior margin, disto-lateral ends lobiform; somite 2 shorter than somite 1, proximal part below carapace posterior margin, disto-lateral ends lobiform; somites 3–5 fused, triangular, proximal one-third of lateral margins convex, middle portion concave, distal one-third narrowed. Pleotelson lingulate, elongated, slightly wider proximally, tapering towards rounded distal end ( Fig. 5 View FIGURE 5 ).
carapace. b–g, holotype male (NMCR-39033): b, carapace; c, right fourth walking leg; d, tip of dactylus of right fourth walking leg; e, left third maxilliped; f, right cheliped; g, various angles of left G1. Scale bars: a, b, d, g and e = 0.5 mm; c and f = 1 mm.
G1 slender, gently twisted at base, distal 2/3 with groove lined with numerous setae, gently curved distally towards midline. G2 short, about half length of G1; tip blunt, slightly expanded laterally.
Colour. This species has completely lost its body pigmentation. However, the sediments from its surrounding environment adhere to their body, giving the animal a yellowish-brown colour.
Etymology. This species, principe , was named after the cave locality, Principe Cave. The name is used as a noun in apposition.
Habitat. Samarplax principe new species, was discovered in Principe Cave, an anchialine cave, located approximately 200 meters away from the shoreline, with the surface salinity ranging from 1.5 to 3.5‰.
Distribution. Samarplax principe new species, is currently known only from Principe Cave, Calicoan Island, Guiuan, Eastern Samar, Philippines.
Remarks. Samarplax principe new species, is the first reported anchialine cave species of Hymenosomatidae exhibiting true troglomorphic characters (see Husana et al. 2009). Ng (1991) described Cancrocaeca xenomorpha , the first true cavernicolous hymenosomatid crab but from a completely freshwater cave system in Sulewesi, Indonesia. The colour of C. xenomorpha in life has been described as ranging from pale yellow to brown (Ng 1991; Deharveng et al. 2002), while S. principe new species, is totally colorless although it appears yellowish-brown owing its coloration to the background sediments that adhere to its body surface.
Non-sexual features of female S. principe new species, generally agree with the males, but in general, they are larger than males. The abdomen of females is dome-shaped and forms a brood cavity that covers the entire thoracic sternum reaching up to the base of the legs. The thoracic sternum has a longitudinal median groove and numerous short setae that are scattered over the surface. In young non-brooding females, the sternum is slightly narrower. The vulvae are oval in shape, sternal in position, elevated and relatively large.
Eggs. A small number of large-sized oval eggs (only 2– 7 eggs measuring about 0.96 x 0.76 mm) is present in the female brood cavity. The small diameter of the female vulvae (about 0.32 x 0.21 mm), however, will not allow the eggs to pass through that opening. This suggests that S. principe new species, may undergo ovoviviparous development similar to euryhaline hymenosomatids like Neorhynchoplax mangalis Ng, 1988 (cf. Ng & Chuang 1996: 59, fig. 26) and N. yaeyamaensis Naruse, Shokita & Kawahara, 2005 (cf. Naruse et al. 2005) (see also Naruse et al. 2008). Furthermore, the small number of eggs of this species may suggest a direct or semi-abbreviated development (P.K.L. Ng, personal communication).
Remarks. Many cave animals have evolved similar features as a result of their adaptation to the cave habitat, like the loss of eyes and possessing long legs (see Culver & Fong 1986; Coineau 2000). The brush-like setae along the inner margin of palm and finger in the chelipeds (instead of cutting teeth) of Samarplax principe new species, is an unusual character, suggesting that it uses its cheliped to brush/scrape off objects to obtain food such as epilithic biofilm (P.K.L. Ng, personal communication). Takeda & Murai (2003) have suggested that fiddler crab Uca panamensis Stimpson, 1859 (Ocypodidae) , uses such a structure to scrape off particles from the rock surface. Although the feeding habit of this species has not been observed, this structure could also possibly function as has been reported for the cavernicolous shrimp Typhlatya mitchelli Hobbs & Hobbs, 1976 , and other atyids. Pohlman et al. (2000) observed that the tufts of setae on the distal end of first and second pereopods of Typhlatya are utilized for grooming and capturing particles from the water. Such setae can be effective because they increase the efficiency in foraging for minute food particles in a food-limited and dark cave environment, and it is not surprising that both the shrimp and crab possess this feature (see also Culver & Fong 1986). Other attributes possessed by cave-adapted crustaceans include the retention of young in the brood cavity to prevent them from drifting too far (see Parzefall 1986) and/or having large but fewer eggs which have energy-richer yolk to help the larvae develop in a more advanced state (see Hüppop 2000). Such life-strategies are useful in a food-limited environment because it increases the survival chances of the young ( Hüppop 2000).
Conservation. Many members of the family Hymenosomatidae are small in size, cryptic in habits and known to inhabit littoral to sublittoral zones that are sensitive to human activities ( Chuang & Ng 1994; Guinot 2011). In the case of Samarplax principe new species, the conservation challenges are associated with the cave environment. In the Philippines, land conversion, development and other associated human activities cause pollution as well as pose problems and threats to natural cave habitats, especially along coastal areas. Clements et al. (2006; 2008) commented that karst areas are rich in biodiversity with high endemism but are threatened by many anthropogenic disturbances. The apparent rarity of the new species described here, its low natality, restriction to the cave habitat and endemism to the island poses an immediate conservation concern.


| ZRC |
Zoological Reference Collection, National University of Singapore |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Brachyura |
|
Family |
|
|
Genus |