
Parartemia yarleensis, Timms, Brian V & Hudson, Peter, 2009
|
publication ID |
https://doi.org/10.5281/zenodo.190741 |
|
DOI |
https://doi.org/10.5281/zenodo.6225435 |
|
persistent identifier |
https://treatment.plazi.org/id/03C38793-FFB2-8150-7BDA-395DFDFF9A1E |
|
treatment provided by |
Plazi |
|
scientific name |
Parartemia yarleensis |
| status |
sp. nov. |
Parartemia yarleensis View in CoL n. sp.
(Figure 7)
Type material. Holotype. Male, SOUTH AUSTRALIA, Yarle Lake system, most likely Choolalie Lake ( 30o17’20”S, 131o31’00”E), (approx 15 km S of Maralinga), 16 September 1979, J. Glover, SAM C6779; Allotype. Female, same collecting data as holotype, SAM 6782; Paratypes. Two males and two females, same collecting data as holotype, SAM C6781.
Other material. About 20 males and five females, SOUTH AUSTRALIA, Yarle Lakes, most likely Choolalie Lake ( 30o17’20”S, 131o31’00”E), (approx 15 km S of Maralinga), 16 September 1979, J. Glover, SAM C6782; many males, SOUTH AUSTRALIA, Lake Labyrinth, ( 30°41’30”S, 135°11’55”E), (approx 27 km NW Kingoonya), 12 June 2004, P. Hudson & G. Tomlinson, SAM C6783; many juveniles, SOUTH AUSTRALIA, “Carters Well Lake”, ( 30°51’01”S, 134°58’35”E), (approx 42 km ESE of Tarcoola), 13 June 2004, P. Hudson & G. Tomlinson, SAM C6808; many juveniles, SOUTH AUSTRALIA, Lake Harris, ( 31o 08’51”S, 135o18’30”E), (approx 20 km S Kingoonya), 19 March 2003, P. Hudson and G. Tomlinson, SAM C6786; many juveniles; SOUTH AUSTRALIA, Ironstone Lagoon, ( 31°42’S, 137°13’30’’E), (approx 65 km SE of Woomera), 1 February, 2007, P. Hudson & G. Tomlinson, SAM C6784; many juvenile males, SOUTH AUSTRALIA, Lake Gilles, ( 33°01’25”S, 136°36’07”E), (approx 20 km NE of Kimba), 6 August 2005, P. Hudson, SAM C6785.
Description. Male. Length 18 mm (head plus thorax 7.5 mm, abdomen 10.5 mm).
Head (Fig. 7A) with first antenna filiform, a little longer than eye plus peduncle. Proximal antennomeres of second antenna fused basially at an angle of about 75 degrees from body axis. Ventral edge of fused antennomeres with paired ventral processes (VP, Fig 7A) three times longer than deep and with length of lateral edge about half that of medial edge. Lateral corner of ventral process protruding slightly, frontal edge only slightly concave, medial corner rounded, and all edges with a few denticles. Small conical mound (CM, Fig, 7B) on ventroposterior surface under lateral corner. Area between ventral processes trapezoid, with a short digitiform medial process (MP, Fig 7A) less than one quarter of depth of medial edge of transverse process. Anterior surface of fused antennomere with paired ridges parallel to body axis and terminating in anterior processes (AP, Fig 7A) with a broad base but digitiform apical half. Anterioventral surface of fused antennomere marked in sunken polygons (SP, Fig 7A). Distal second antenna antennomere subcylindrical, slightly concavely curved and tapering to a sharp apex. Length about 1.6 times proximal antennomere. Labrum without a spine.
Thorax gradually widening posteriorly to 11th segment, mainly by increasingly larger lateral lobes, tending asymmetrical (i.e. maximum width displaced from middle of lobe) segments 7–10 (L, Fig. 7C). Genital segments narrower than 11th thoracic segment and abdomen continually narrowing so that 6th segment about 2/ 3rds width of first segment.
Thoracopods (Fig 7E) of the Parartemia type as described for P. acidiphila n. sp., except for fewer posterior setae on endites 1+2, 3, endopodite and exopodite (c.45, 11, 26 and 35 respectively). Posterior setae on medial edge of endopodite more numerous than usual (11 cf c. 7), particularly strong, curved apically and with a short pecten apically (EPS, Fig 7E).
Paired gonopods with a spines subapically and a short digitiform processes (DP, Fig. 7D) on the shoulder of wider basal part, neither hooked. No type specimens with gonopods everted.
Abdominal segments serially decreasing in diameter and increasing in length posteriorly. Sixth segment about twice as long as first segment. Cercopods subequal in length to sixth abdominal segment and with setae medially and laterally.
Description. Female. Length 11.3 mm.
Head (Fig. 7F) with first antenna filiform, about length of eye plus peduncle. Second antenna about twice length of eye plus peduncle, flattened and with its widest area about two-thirds its length towards the apex, followed by a marked narrowing to an acute apex on the posterior side. Apex curved like the recurved labrum spine.
Thoracic segments (Fig. 7G) expanded laterally by distinct lobes, increasing in size and degree of asymmetry (i.e. displacement of widest point from centre of lobe) serially segments 5 to 9. Segment 10 with very different lateral lobes, expanded anteriorly, free in allotype and dorsal to lobe of 9th segment. Segment 11 with a narrow triangular lateral lobe. In lateral profile (Fig. 7H), segments 9 to 11 not raised dorsally as much as anterior segments. Segment 8 swollen dorsally. Paired brood pouches separate, oval, unlobed but joined ventrally to a gonoduct shorter than the depth of the brood pouch. Each pouch with numerous spherical smooth surfaced eggs.
Thorax with only 10 pairs of thoracopods and 10th thoracopod reduced to about half size of other thoracopods. Anterior setae of 10th thoracopod typical, but few posterior setae on all parts and lacking an epipodite but with reduced praepipodite (Fig. 7I). Fifth thoracopod as in male.
Abdomen as in male, but surface denticulate.
Etymology. The species is named after the type locality.
Variability. Though this species is known from a few sites, many of these had only juvenile males, so variation between sites is hardly studied. Within the type locality some males had more bulbous lateral corners to their ventral processes than the holotype. Among females, the second antennal apex is not always curved and the lateral lobe of the 10th thoracic segment varied within and between sites, with it being attached to the lobe of the 9th segment, often in younger females.
Differential diagnosis. Male P. yarleensis has a head (specifically a medial process and ventral processes) broadly similar to those of P. informis , P. serventyi and P. contracta , but unlike those species, has distinct thoracic lobes. These lobes are not as large as in P. cylindrifera and in Parartem ia sp. g (as illustrated in Timms 2004), but broadly similar to those of P. auriciforma n. sp. and P. triquetra n. sp. While the latter two species occur in the same general area as P. yarleensis n. sp., they are easily distinguished as neither have the medial process between the ventral processes as in P. yarleensis n. sp.
Female P. y a r l e e n s i s n. sp. also share many features with local species P. auriciforma n. sp. and P. triquetra n. sp., such as thoracic lateral lobes, round to oval brood chambers and greatly reduced or absent 11th thoracopods. However, P. yarleensis is distinctive by reason of superficial dorsal swelling on the 8th segment (somewhat like that in P. serventyi ), and the bulbous lateral lobes of segment 10. It cannot be confused with P. serventyi as this species has posterior lobes on its brood pouches, no thoracic lateral lobes, and paired dorsolateral swellings on segment 9.
Type locality. Yarle Lakes is a series of lakes south of Maralinga that fill episodically and are ‘very saline’ according to collecting data. There is some doubt over which one of the lakes was sampled, but examination of the field notes of J. Glover’s participants of the field trip, the probable collection site of Choolalie Lake was established on the basis of sketch map of the area.
FIGURE 7. Parartemia yarleensis n. sp. Male A-E, Holotype; Female, F-I Allotype; both from Yarle Lakes, most likely Lake Choolalie, SA. A, anterior view of head with first and second antennae (VP = ventral processes, AP = anterior processes, MP = medial process, SP = sunken polygons); B, posterior view of one side of basal antennomere of second antenna showing the conical mound (CM) behind the ventral process; C, dorsal view of body from head to cercopods showing segmental lobes (L); D, gonopods with genital segments showing digitform processes (DP); E, Fifth thoracopod with pectin bearing endopod posterior setae labelled (EPS); F, Lateral view of head; G, dorsal view of thoracic segments 4-11, genital segments, brood pouches and first two abdominal segments; H, lateral view of posterior thorax and adjacent brood pouch; I, 10th thoracopod. Scale bars 1 mm.
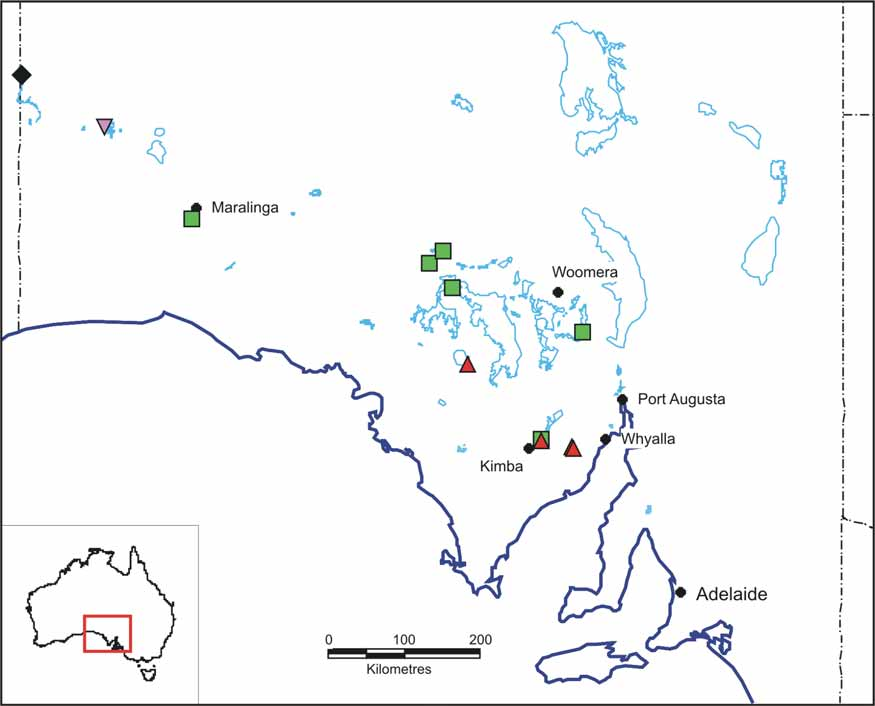
Distribution and ecology. P. yarleensis n. sp. is known from a broad arc of lakes extending from Woomera to Maralinga in the northwest of South Australia ( Fig. 4 View FIGURE 4 ). The collection from Ironstone Lagoon also contains P. m i n u t a, a species smaller than P. y a r l e e n s i s ( P. m i n u t a males mean 8.7 mm, 10 specimens, females mean 5.2 mm, 10 specimens). Such a congeneric occurrence is rare in Parartemia (A. Savage, pers. comm.; B. Timms, unpublished data) possibly because most species of Parartemia do what P. zietziana does and that is they live on resuspended organic matter ( Marchant and Williams, 1977), as opposed to algal eating Branchinella , in which congeneric occurrence of different sized species and hence filtering ranges, are common (e.g. Timms & Sanders, 2002).
| SAM |
South African Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |