
Grubeulepis serrata, Cutrim & Praseres & Conceição & Almeida & Júnior & Oliveira, 2018
|
publication ID |
https://doi.org/10.11646/zootaxa.4441.2.11 |
|
publication LSID |
lsid:zoobank.org:pub:4D687F78-ED40-4FE1-898E-C7ADEA957A1D |
|
DOI |
https://doi.org/10.5281/zenodo.5978163 |
|
persistent identifier |
https://treatment.plazi.org/id/03C387D6-4358-723D-FF03-FE928805FD7D |
|
treatment provided by |
Plazi |
|
scientific name |
Grubeulepis serrata |
| status |
sp. nov. |
Grubeulepis serrata View in CoL sp. nov.
Figures 2–7 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 6 View FIGURE 7
Material examined. Holotype: Pucal Beach, Raposa, Maranhão, Brazil ( 02°25’33.4” S 44°07’25.8” W), 12 February 2016 (CIPY-1737). GoogleMaps
Paratypes: 0 9 paratypes. Pucal Beach, Raposa, Maranhão, Brazil, 02°25’33.4” S 044°07’25.8” W: 12 February 2016 ( 1 paratype, CIPY –1741); 20 June 2016 ( 2 paratypes, CIPY –1742); 22 August 2016 ( 1 paratype, CIPY –1744); 19 October 2016 ( 1 paratype, CIPY –1738); 19 October 2016 ( 2 paratypes, CIPY –1740); 13 December 2016, ( 1 paratype, CIPY –1739); 13 December 2016, ( 1 paratype, CIPY –1743).
Diagnosis. 1 pair of eyes, 14 pairs of branchiae, posterior lamellae beginning in the segment 27, acicular neurochaeta with serrated outer edge present in the 3rd segment.
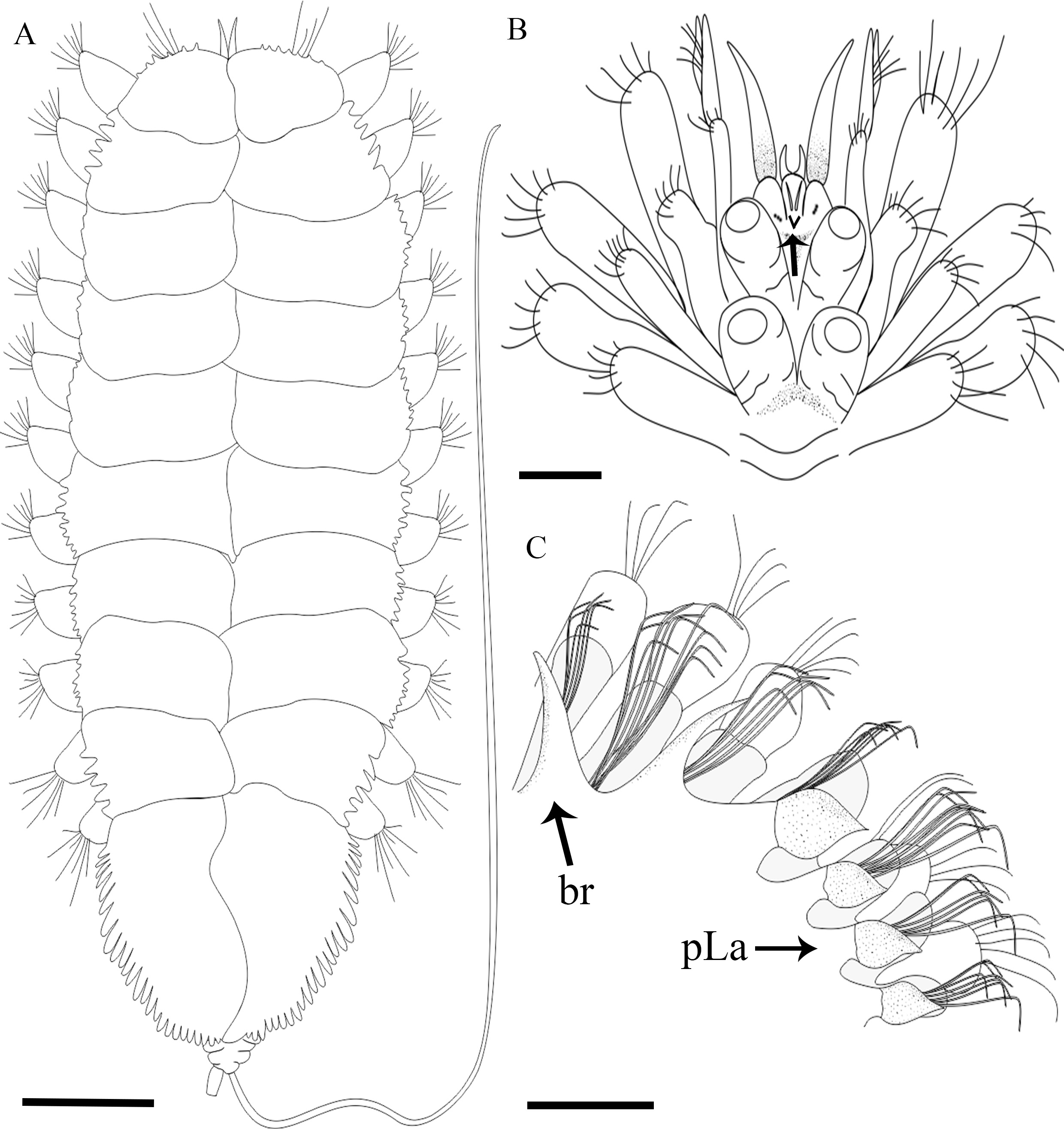
Description. The holotype is a complete specimen with 33 segments, 8.0 mm in length and 2.0 mm wide on the median part of the body ( Fig. 2A View FIGURE 2 ). Prostomium covered by the second segment, median antenna short and conical, with dorso-lateral insertion; lateral antennae conical with ventral insertion, relatively long palps and extended between the tentacular cirri ( Fig. 2B–C View FIGURE 2 ); one pair of eyes. 12 pairs of elytra with lateral processes and digitiform articulations, first pair of elytra with 14 papillae in the anterior margin, second pair with seven lateral processes bi or triarticulated, eighth pair with 13 lateral processes bi or triarticulated, twelfth pair of elytra with 25 lateral processes biarticulated ( Fig. 4A–D View FIGURE 4 ). 14 pairs of branchiae from the 7th segment, projected dorsal to the parapodia and defined by the presence of branchial cirri. From segment 27, branchiae are substituted by the foliaceous lamellae which decrease in size until the end of the body ( Fig. 2D View FIGURE 2 ). Parapodia biramous ( Fig. 3A–D View FIGURE 3 ). Neurochaeta acicular with serrated outer edge present in the 3rd segment ( Fig. 3A View FIGURE 3 , 5E). Notochaetae of several types along body: i) curved with distal end slightly serrated dorsally, spoon-shaped distal and hooked upper notochaetae (Fig. 5C–D, 6C); ii) smooth with thin distal end and capillaries with denticles (Fig. 5B). Posterior notochaetae thicker than anterior ones, curved, serrated in the distal region and with thin distal end ( Figure 6D View FIGURE 6 ). Neuropodial pectinate and capillaries in the anterior region (Fig. 5A). Posterior neurochaetae robust and curved, spiny distally and with spatulated or thin distal end ( Fig. 6A–B View FIGURE 6 ). Pygidium with long right-handed anal cirrus finely serrated in one side and with short and round left-handed anal cirrus ( Fig. 7 View FIGURE 7 ).
FIGURE. 5. Grubeulepis serrata sp. nov. A, Neuropodial pectinate and capillaries chaeta; B, Notopodial capillaries chaetae with denticles; C–D, Hooked upper notochaetae (arrow); E, Acicular neurochaeta of the parapodium 3. Scales: A–C = 0.5 mm; D = 0.03 mm; E = 0.05 mm.
Morphological variations: Paratypes with 21 to 39 segments, with body length and width ranging from 3.0 to 10.0 mm and 0.8 to 2.0 mm respectively. First pair of elytra with 12–14 papillae in the anterior margin; second pair of elytra with 5–7 lateral processes bi or triarticulated, eighth pair with 10–14 lateral processes bi or triarticulated, twelfth pair of elytra with 23–25 lateral processes (in adults) biarticulated. In young individuals the elytra cover the whole body and the twelfth pair is smaller than the others and bears 12–13 lateral processes. 13–14 pairs of branchiae in adults and 6–7 in juveniles.
Colour. Fixed individuals without coloration.
Habitat. Sandy beach dominated by very fine sand.
Distribution. Atlantic Ocean, Brazil, São Luís Island, Maranhão.
Etymology. Name derived from the Latin word serrata ( serra), referring to the acicular neurochaeta with serrated outer edge present in the third segment.
Remarks. Grubeulepis serrata sp. nov. can be distinguished from the other 11 known species of the genus by having: one pair of eyes, 14 pairs of branchiae, posterior lamellae beginning in the 27th segment, hooked upper notochaetae with a dorsally serrated end and serrated acicular neurochaeta in the third segment (Table 1). It is noteworthy to emphasize that 14 pairs of branchiae have not been recorded for the genus before and thus the diagnosis of the genus had to be amended (see above). Grubeulepis kurnai ( Woolley &Wilson 2011) also has serrated hooked upper notochaetae, but in this species the part of the distal end is dorsal while in Grubeulepis serrata sp. nov. the part of the distal end is ventral. Moreover, both species differ also in many characters such as the presence of eyes, number of branchiae, number of papillae in the first elytra, and number and articulation of lateral processes of the elytra (Table 1). Among Brazilian species, Grubeulepis serrata sp. nov. shares with G. augeneri and G. fimbriata spoon-shaped notochaeta, but differs from them in the presence of spines, the number of lateral processes of the elytra, pairs of branchiae and eyes. Grubeulepis serrata sp. nov. is similar to G. tebblei regarding the robust neurochaetae with spatulated distal end and the segment of the appearance of the posterior lamellae, however G. tebblei does not have acicular neurochaeta and both species also differ on the number of lateral processes of the elytra, number of branchiae, eyes and the hooked upper notochaetae (Table 1). Grubeulepis serrata sp. nov. resembles G. mexicana by the number of lateral processes in the 12th elytra, the segment where foliaceous lamellae appear, and by the presence of acicular neurochaetae in the third segment, but differ by the serrated outer edge acicular in Grubeulepis serrata sp. nov. (Table 1). Moreover, they also differ in the number of branchiae, eyes and the types of notochaetae and neurochaetae: Grubeulepis serrata sp. nov. has 14 pairs of branchiae, one pair of eyes and notochaetae with spoon-shaped distal end, while G. mexicana has 11 pairs of branchiae, 2–5 pairs of eyes and spiny notochaetae ( Berkeley & Berkeley 1939; Table 1). Grubeulepis serrata sp. nov. shares with G. geayi the spiny notochaetae with spoon-shaped distal end, but differ in the number of lateral processes, the elytra articulations and the segment with the acicular neurochaetae, third in Grubeulepis serrata sp. nov. and third and fourth in G. geayi ( Fauvel 1918; Table 1).
The eyes of these worms are not necessarily a reliable character since they may present high intraspecific variability (e.g. Pettibone 1986). Yet, all the 10 individuals studied here of Grubeulepis serrata sp. nov. had a single pair. Additionally, the eyes are small and commonly hidden by the second chaetiger and thus easily overlooked, particularly when the prostomium is pigmented ( Pettibone 1969). Thus, studies on the group should take particular care on this issue.
The genus Grubeulepis commonly has morphological variations through its ontogenetic development, such as reported here. Young Grubeulepis have the elytra covering the entire body and may easily be confused with Mexieulepis , distinguished by the presence of acicular neurochaetae. This chaetal type appear in few chaetigers or are absent in Grubeulepis , but occur in most chaetigers in Mexieulepis ( Uebelacker 1984; Pettibone 1986). The taxonomic use of the number of lateral processes and articulation of the elytra demands caution since they are highly variable, commonly with juveniles differing markedly from adults as observed here for Grubeulepis serrata sp. nov. For instance, young G. mexicana have articulated lateral processes while adults do not ( Pettibone 1986), and in G. kurnai articulated lateral processes are substituted by non-articulated during growth ( Woolley & Wilson 2011). These observations emphasize the need to characterize individuals of different developmental stages in order to more precisely delimitate the species and avoid taxonomic uncertainties and/or misidentifications, helping to understanding variability in these worms.
TABLE]. Comparison of main characteristics of Grubeulepis species known worlđwiđe (mođifieđ from Wooley & Wilson 2011).
……continued on the next page
TABLE]. (Continueđ)




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |