
Trechus eoanophthalmus, Schmidt & Hoffmann & Michalik, 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4083.3.7 |
|
publication LSID |
lsid:zoobank.org:pub:C999EBFD-4EAF-44E1-B7E9-95C9C63E556B |
|
DOI |
https://doi.org/10.5281/zenodo.6067615 |
|
persistent identifier |
https://treatment.plazi.org/id/03C387EA-101B-3610-FF2B-DC47FC611B3D |
|
treatment provided by |
Plazi |
|
scientific name |
Trechus eoanophthalmus |
| status |
sp. nov. |
Trechus eoanophthalmus View in CoL sp. n.
Figs. 1–13 View FIGURES 1 – 2 View FIGURES 3 – 4 View FIGURES 5 – 7 View FIGURES 8 – 12 View FIGURE 13
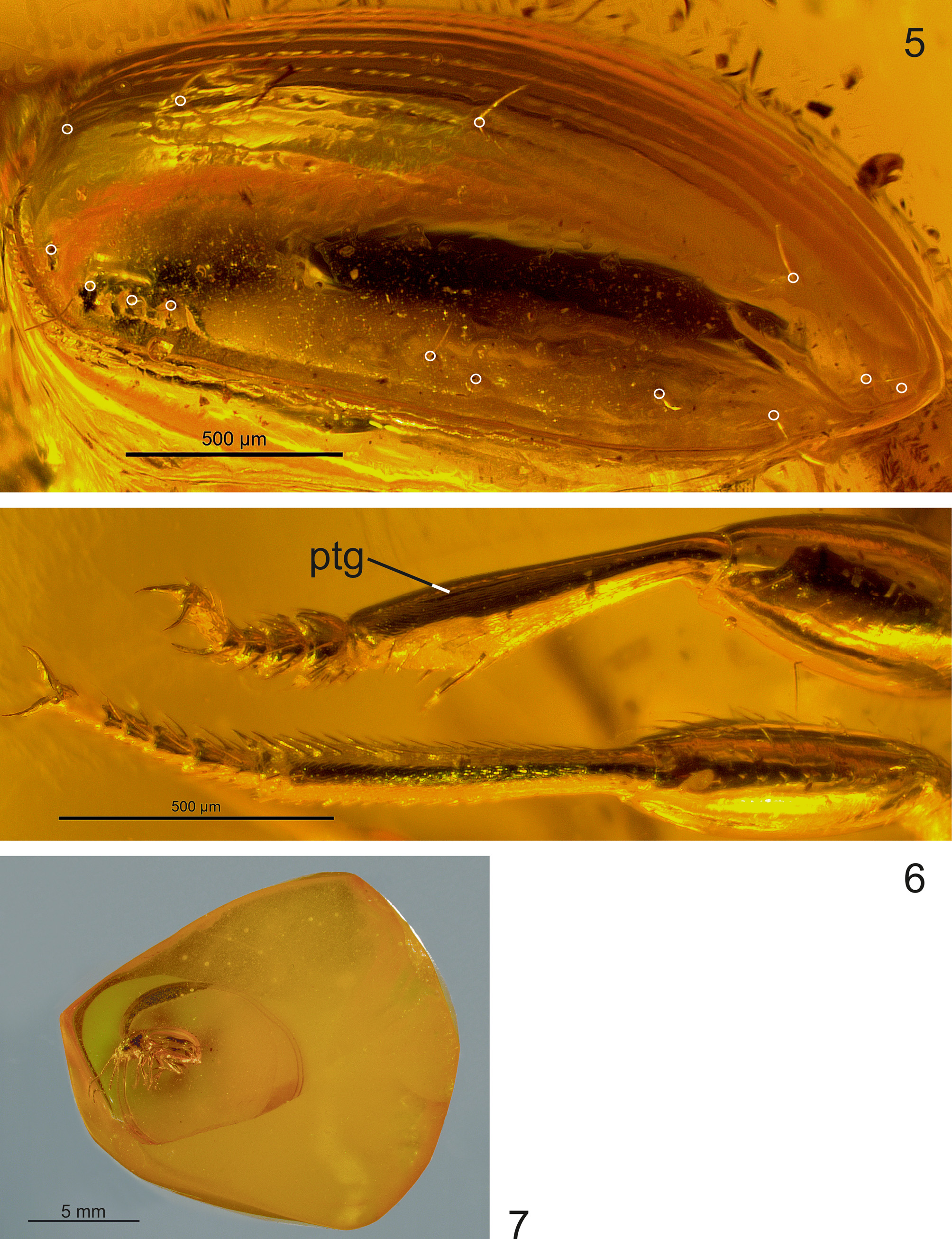
Holotype. Male in Baltic amber; size of piece approximately 17.0 x 15.6 x 8.6 mm (irregularly cut, Fig. 7 View FIGURES 5 – 7 ), No. 452 in the collection F. Eichmann, Hannover, Germany (likely to be later deposited at the Geologisch- Paläontologisches Museum, University of Hamburg, Germany).
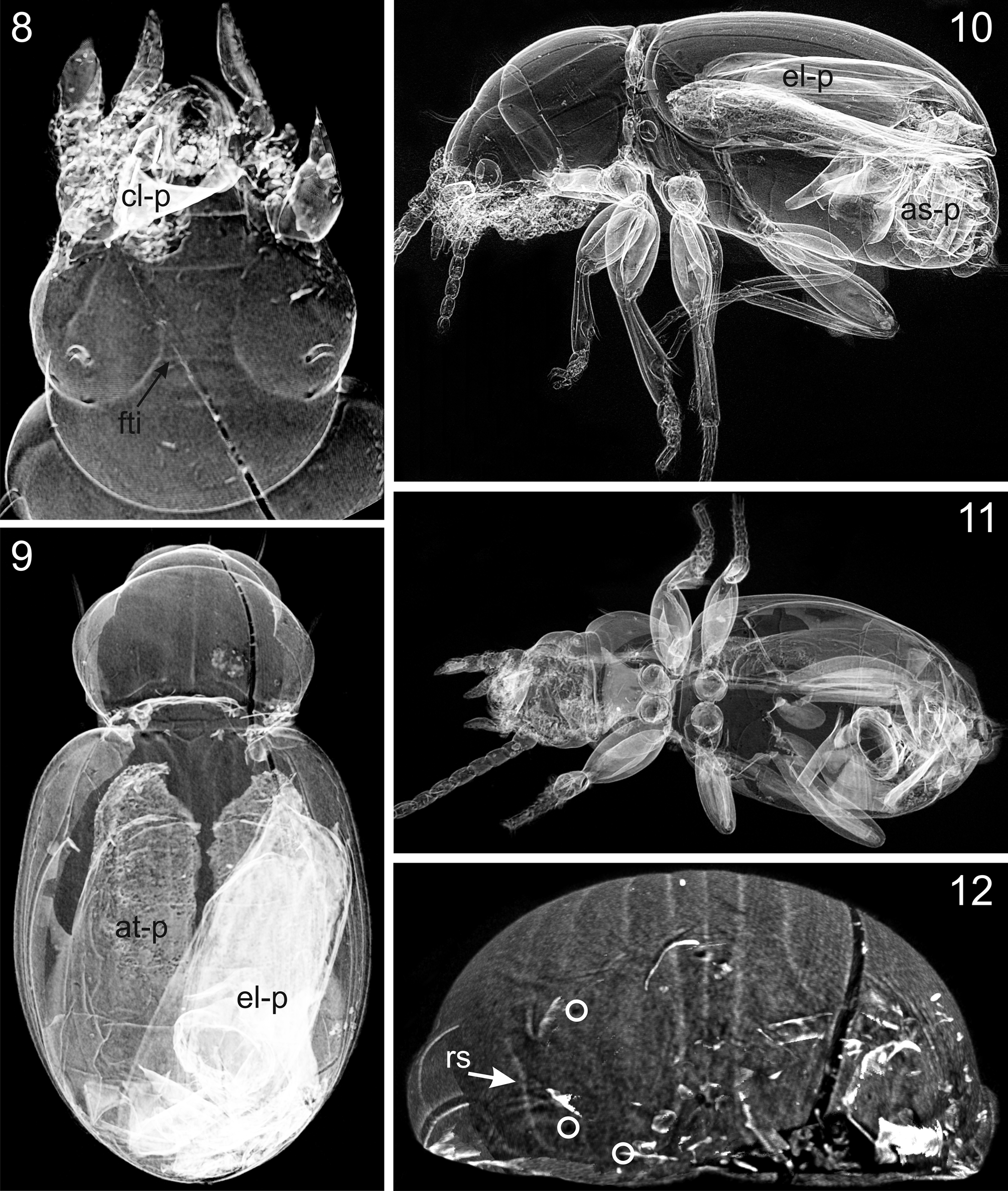
Preservation status. The specimen is markedly shrunken with its exoskeleton, which is completely dissociated from the inclusion wall. The latter represents the negative imprint of the beetle body and is visible in most portions. Detached parts of the original beetle exoskeleton, e.g., the distorted right elytron, which moves freely in the cavity of the amber piece, and the shrunken legs, are visible under optical microscope and could be imaged in detail using micro-CT ( Figs 8–11 View FIGURES 8 – 12 ). This particular condition of preservation is probably the result of the embedding of an immature and thus teneral specimen whose exoskeleton collapsed after the resin was cured.
Most parts of the negative imprint of the beetle body were visible in detail using a light microscope (see, e.g., microsculpture of head, Fig. 3 View FIGURES 3 – 4 , protibial groove, Fig. 6 View FIGURES 5 – 7 ). However, since the amber piece was polished only from two sides lateral to the inclusion ( Fig. 7 View FIGURES 5 – 7 ) a view directly from dorsad and ventrad of the beetle is not possible due to refractions. The mouth parts are surrounded by dirt particles and therefore, the basal portion of mandibles, maxillae and mentum could not be examined using light microscopy (only the apical part of mandibles and apical segments of the palpi were visible, Figs. 3 View FIGURES 3 – 4 ). In addition, the results of micro-CT imaging showed that the imprint of the inclusion was destroyed on the ventral surface of head ( Fig. 11 View FIGURES 8 – 12 ) and therefore, diagnostic characters of the mentum could not be reconstructed.
Syninclusions. Apart from several dirt particles, no syninclusions are preserved in the amber piece.
Description. Body length: 4.4 mm.
Colour: Colour of body surface undeterminable.
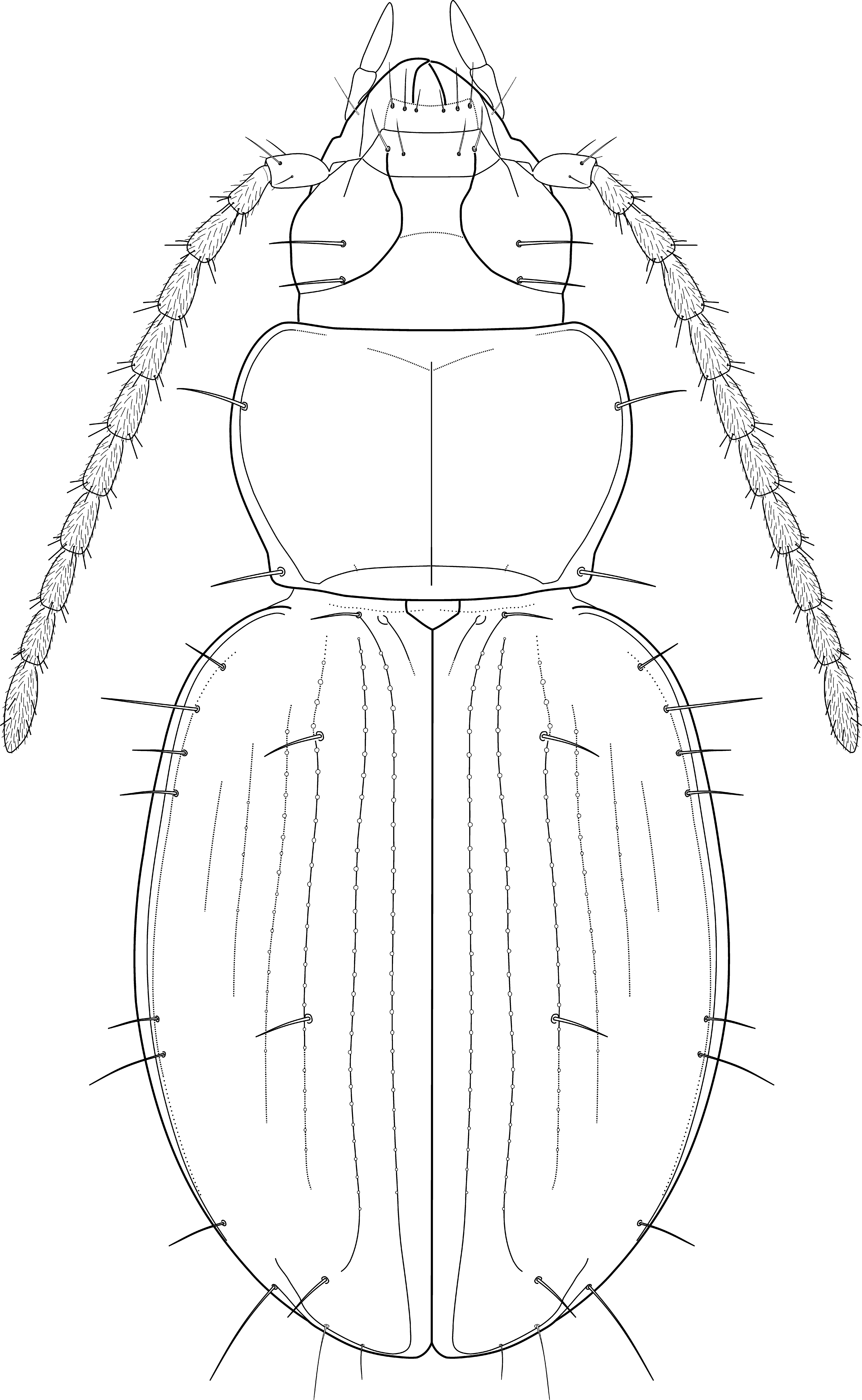
Microsculpture: Clypeus and frons with slightly engraved markedly transverse meshes, neck with larger and less transverse meshes ( Fig. 3 View FIGURES 3 – 4 ). Discs of pronotum and elytra with very fine and weakly impressed transverse meshes (visible under magnification> 150 x), appearing polished under lower magnifications.
Head: Length 1.09 mm, short and robust, notably broad near base ( Fig. 8 View FIGURES 8 – 12 ). Mandibles of normal length for Trechus (mandibular dentition and chaetotaxy of the mandibular groove could not be reconstructed). Labrum with three pairs of setae (shape of the labral apical margin could not be displayed). Clypeus with two pairs of setae in normal position. Shape of apical segments of maxillary and labial palpi as in Trechus sensu stricto. Eyes completely reduced ( Figs 1–4 View FIGURES 1 – 2 View FIGURES 3 – 4 , 8 View FIGURES 8 – 12 ).Tempora, frons, orbital and supraorbital areas smooth, markedly convex, with supraorbital furrows deep, complete, and almost uniformly bent on disc ( Figs 3–4 View FIGURES 3 – 4 ); frons with a slight but distinct transverse impression ( Figs 3 View FIGURES 3 – 4 , 8 View FIGURES 8 – 12 ); two supraorbital setae present and in normal position for Trechus . Antennae moderately short, with scapus rather robust (length 0.17 mm; length/width = 1.31; 1.3times broader than pedicellus), and with pedicellus and third antennomere of the same length (0.16 mm) and shape.
Prothorax: Pronotum markedly transverse (width/length = 1.54), length 0.82 mm, 1.39 times broader than head, broadest in middle, with sides evenly rounded in anterior 5/6 and straight before the small but well developed laterobasal angles; basal margin 1.25 times broader than apical margin. Disk markedly convex, smooth ( Fig. 4 View FIGURES 3 – 4 ). Anterior margin almost straight, very finely beaded in outer third, smooth in middle, with apical angles very slightly protruded, rounded. Posterior margin not beaded, moderately convex in middle, slightly incised before outer quarters; laterobasal angles small, not shifted anteriad, almost rectangular, not protruded laterally. Median longitudinal impression distinct, somewhat deeper near base, disappearing near apex; anterior transverse impression very fine, smooth; posterior transverse impression deep and linear, smooth, laterobasal foveae indistinct; pronotal base convex between laterobasal foveae and side margin. Lateral gutter moderately narrow throughout, very slightly widened towards base, smooth. Both lateral and laterobasal setae present, with the lateral seta located at apical third of pronotum. Proepisternum and prosternal process glabrous and smooth, latter without margin.
Pterothorax: Elytra markedly convex on disc ( Figs. 1–2 View FIGURES 1 – 2 , 10 View FIGURES 8 – 12 ), in dorsal view short-ovate ( Fig. 9 View FIGURES 8 – 12 ), length 2.49 mm, length/width = 1.21, widest near their mid-length, moderately wider than pronotum (width of elytra/width of pronotum = 1.20), glabrous beside normal setation. Humeral angles fully rounded, basal groove absent. Parascutellar stria and parascutellar seta present. First stria complete, second stria disappearing near apex, third and fourth striae distinctly finer impressed than inner striae, disappearing near base and apex, fifth and sixth striae indistinct, seventh and eighth striae absent; inner striae finely punctated apart from the apical fifth of elytra ( Figs. 5 View FIGURES 5 – 7 , 13 View FIGURE 13 ). Each elytron with two discal setae in third stria, and with preapical seta, situated at apical eighth of elytra, almost equally as close to the apical margin as to the suture of elytra ( Figs 5 View FIGURES 5 – 7 , 12 View FIGURES 8 – 12 ). Recurrent stria extending towards the reduced fifth stria anteriorly, moderately long, almost straight, with front end situated slightly anterior of the level of the preapical seta ( Fig. 12 View FIGURES 8 – 12 ). Setae of umbilicate humeral series close to the elytral margin, with posterior three setae equidistant from each other, and with first seta slightly more distant from second ( Figs. 5 View FIGURES 5 – 7 , 13 View FIGURE 13 ). The two setae of the medial group of the umbilicate series far removed from the preapical group ( Figs. 5 View FIGURES 5 – 7 , 13 View FIGURE 13 ). Epipleura long, nearly reaching the apex. Metepisternum short, glabrous and smooth, with outer margin 1.2 times longer than anterior margin.
Abdomen: Abdominal sternites V–VII and anal sternite each with one (male) pair of setae near apical margin; surfaces smooth, without hairs or micropunctures.
Legs: Moderately short, robust, all femora unmodified; protibia straight and moderately dilated towards apex, glabrous on their anterior surface, with a short and very fine longitudinal groove on external surface ( Fig. 6 View FIGURES 5 – 7 ). Basal two protarsomeres moderately dilated ( Fig. 6 View FIGURES 5 – 7 ).
Male genitalia: Could not be located within the collapsed abdominal portion of the exoskeleton.
Etymology. The name of the new species is a combination of “eo“, derived from the Eocene epoch when the beetle was fossilized, and “anophthalmus” (= eyeless), derived from a combination of the Greek words “a(n)” = without, and “ophthalmos” = eye.
Relationships. Trechus eoanophthalmus sp. n. shares all diagnostic features with the previously described T. balticus placing the new fossil species either in the main Trechus clade sensu Faille et al. (2013) or in the Nearctic lineage Microtrechus Jeannel, 1927 (see discussion in Schmidt & Faille 2015): shape of mandibles normal; orbital area glabrous, with deep furrows semicircularly separating well developed supraorbital region from frons and neck; two supraorbital setae present; apical segment of maxillary palpus as long as the penultimate, at base almost as broad as apex of penultimate segment, and at apex regularly conically pointed; pronotum and elytra glabrous; pronotum with posterior transverse impression and laterobasal foveae linear and sharply limited towards the convex disc, with one anterolateral and one laterobasal setae each side; elytra with basal groove absent, and with two discal setae; recurrent sutural stria forming a deep carina directed towards the end of the fifth stria; apical triangle present, setae of humeral group situated close to the elytral margin, setae of medial group well separated from the preapical group; apical portion of protibia glabrous on external surface.
Trechus eoanophthalmus View in CoL sp. n. is distinguished from Microtrechus by two dilated male protarsomeres. Thus, the new species is most likely a representative of the main Trechus View in CoL clade sensu Faille et al. (2013). However, phylogenetic studies have shown that the genus Trechus View in CoL sensu auctorum is a non-monophyletic assemblage (see Faille et al. 2010, 2011, 2013). In addition, the phylogenetic positions of many of the non-European lineages need still to be investigated. Like in the previously described Baltic amber fossil T. balticus View in CoL , phylogenetically important genital characters could not be revealed in T. eoanophthalmus View in CoL sp. n. due to the poor preservation of the internal parts of its body. Consequently, the systematic position of T. eoanophthalmus View in CoL sp.n. within the mega-diverse genus Trechus View in CoL remains dubious. Nevertheless, it seems very likely that the Eocene fossil species, T. balticus View in CoL and T. eoanophthalmus View in CoL sp.n., are closely related and may thus represent members of the same (extinct?) branch within Trechus View in CoL sensu lato. This hypothesis is based on two derived character states, which are potential synapomorphies of the two fossil species: i) microsculpture of pronotum and elytra consist of very fine very weakly impressed transverse meshes; ii) elytral striation markedly reduced laterally and apically. However, since these character states are also present in some of the extant Trechus View in CoL species, this hypothesis needs further testing based on a more extensive fossil data set.
A particular character of the fossil T. eoanophthalmus View in CoL sp.n. is the complete reduction of the compound eyes. Within the extant world fauna of Trechini View in CoL a huge number of anophthalmous and microphthalmous species are described, and their diversity is particularly rich in the southern parts of the Western Palaearctic (e.g., Jeannel 1928, Casale & Laneyrie 1982, Casale et al. 1998). It has been shown, however, that different degrees of eye reduction evolved several times independently at least within Trechus View in CoL sensu lato ( Casale 2011, Faille et. al. 2010, 2011, 2013, 2014). Therefore, “eyelessness” as present in the fossil T. eoanophthalmus View in CoL sp.n. is likely not an indication for a close relationship with any of the extant eyeless Trechini View in CoL lineages.
Differential diagnosis. The Baltic amber fossil species Trechus balticus is the only trechine beetle known from Paleogene deposits to date ( Schmidt & Faille 2015). Trechus eoanophthalmus sp.n. is easily distinguished from the latter by the completely reduced eyes, larger and more robust body, broader lateral gutter of pronotum, and the presence of a fine groove on external surface of the protibia. Trechoides fasciatus Motschulsky, 1856 is another Baltic amber fossil with uncertain taxonomic position within Carabidae , and its holotype specimen was very probably lost ( Schmidt & Faille 2015). Comparison has thus to be based on the few characters given in the original description. Trechus eoanophthalmus sp.n. is distinguished from the latter taxon by the much larger body size (almost three times longer), the reduced eyes, and the apically rounded elytra.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Tribe |
Trechini |
|
Genus |
Trechus eoanophthalmus
| Schmidt, Joachim, Hoffmann, Hannes & Michalik, Peter 2016 |
Trechus eoanophthalmus
| Schmidt & Hoffmann & Michalik 2016 |
Trechus
| Schmidt & Hoffmann & Michalik 2016 |
Trechus
| Schmidt & Hoffmann & Michalik 2016 |
T. eoanophthalmus
| Schmidt & Hoffmann & Michalik 2016 |
T. eoanophthalmus
| Schmidt & Hoffmann & Michalik 2016 |
Trechus
| Schmidt & Hoffmann & Michalik 2016 |
T. eoanophthalmus
| Schmidt & Hoffmann & Michalik 2016 |
Trechus
| Schmidt & Hoffmann & Michalik 2016 |
Trechus
| Schmidt & Hoffmann & Michalik 2016 |
T. eoanophthalmus
| Schmidt & Hoffmann & Michalik 2016 |
Trechus
| Schmidt & Hoffmann & Michalik 2016 |
T. eoanophthalmus
| Schmidt & Hoffmann & Michalik 2016 |
T. balticus
| Schmidt & Faille 2015 |
T. balticus
| Schmidt & Faille 2015 |
Microtrechus
| Jeannel 1927 |
Trechini
| Bonelli 1810 |
Trechini
| Bonelli 1810 |