
Coluber, Linnaeus, 1758
|
publication ID |
https://doi.org/10.5252/geodiversitas2020v42a20 |
|
publication LSID |
urn:lsid:zoobank.org:pub:8FF2A078-CE45-4BF1-A681-00136F57375E |
|
DOI |
https://doi.org/10.5281/zenodo.4488227 |
|
persistent identifier |
https://treatment.plazi.org/id/03C587C7-4315-FFF7-FBAB-FA2F4C9BFA7F |
|
treatment provided by |
Felipe |
|
scientific name |
Coluber |
| status |
|
Coluber (s.l.) sp.
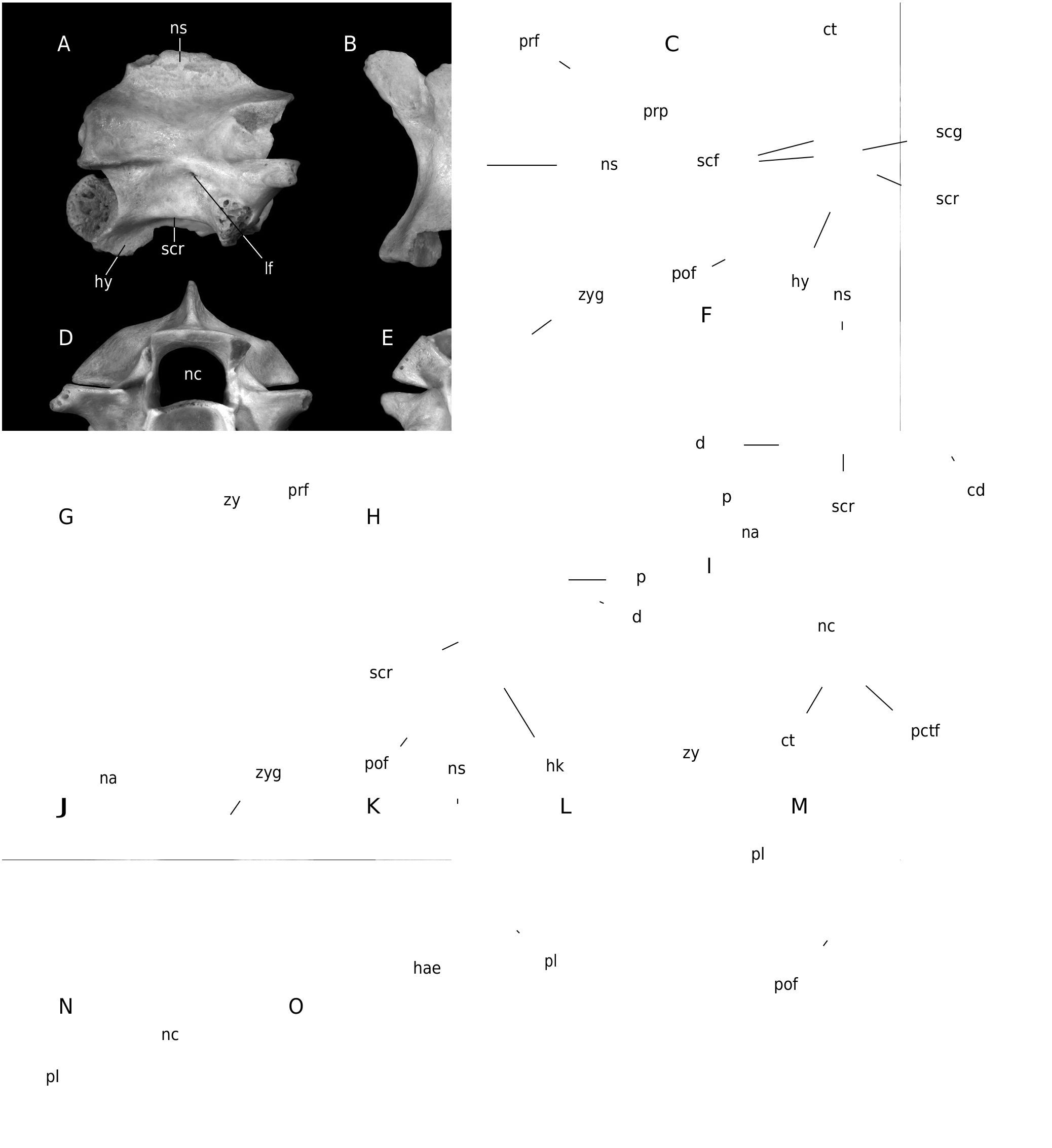
( Fig. 9 View FIG )
Coluber sp. 1 – Ivanov & Musil 2004: 229 (in part).
Coluber sp., type I – Ivanov et al. 2006: 229, table 2 (in part).
MATERIAL. — MWQ, early Miocene, Burdigalian, Orleanian, MN 4: 1/2001 Turtle Joint: Two anterior trunk vertebrae (Pal. 1454-1455), 6 middle trunk vertebrae (Pal. 1456-1461), 2 caudal vertebrae (Pal. 1462, 1463). 2/2003 Reptile Joint:Two trunk vertebrae (Pal. 1956- 1957), 2 caudal vertebrae (Pal. 1958-1959).
DESCRIPTION
Anterior and middle trunk vertebrae
The only two preserved anterior trunk vertebrae differ from those from the middle trunk portion by the presence of hypapophysis instead of haemal keel and higher neural spine (Pal. 1454; Fig. 9 View FIG A-E). All preserved middle trunk vertebrae are at least partially fragmentary with neural spines mostly broken-off close to their bases. In lateral view, the preserved neural spine base of the most complete specimen (Pal. 1456; Fig. 9 View FIG F-J) indicates that the neural spine was probably at least twice longer than high. The caudal margin of the neural spine was inclined posteriorly as documented by specimen Pal. 1956. The well-developed interzygapophyseal ridges are short. The lateral foramina occur just below these ridges. The prominent subcentral ridges are moderately dorsally arched. The large diapophyses are well-separated from somewhat smaller parapophyses. Although parapophyses are incomplete in all specimens, their ventral margin apparently extended below the cotylar rim. In dorsal view, the wide zygosphene has developed lateral lobes. The medial lobe is rather wide. The prezygapophyseal articular facets are oval with long axis directed antero-laterally. The prezygapophyseal processes are broken-off close to their bases. The epizygapophyseal spines are moderately developed. A deep notch occurs at the caudal margin of the neural arch. In ventral view, the straight subcentral ridges form the lateral margins of a cranio-caudally elongated centrum of narrowly triangular shape. The subcentral grooves are wide and shallow. The subcentral foramina are rather small. The haemal keel is narrow and reaches posteriorly almost to the cranial margin of the small rounded condyle. Small subcotylar tubercles occur at the ventral margin of the cotylar rim. The postzygapophyseal articular facets are subrectangular and slightly laterally elongated. In cranial view, the neural arch is vaulted, and the neural canal is wide and rounded with wide and shallow lateral sinuses. The cranial margin of the zygosphenal lip arches dorsally. The small paracotylar foramina occur within depressions on either side of the rounded cotyle. In caudal view, the zygantrum is wide. The vertebral dimensions of the largest specimen (Pal. 1456) are as follows: cl = 4.52 mm; naw = 3.21 mm; cl/naw = 1.41.
Caudal vertebrae
The rarely preserved caudal vertebrae are relatively short. In lateral view, the neural spine of the better-preserved specimen was twice as long as high. Its cranial margin rises in the middle of the zygosphene length. The zygosphenal facets are oval. The preserved base of the right pleurapophysis is directed antero-ventrally. Haemapophyses are broken-off close to their bases. In dorsal view, the zygosphene is almost straight. The prezygapophyseal articular facets are oval in outline. The preserved base of the right pleurapophysis in specimen Pal. 1462 ( Fig. 9 View FIG K-O) is anterolaterally directed indicating anterior caudal position within the vertebral column. In cranial view, the zygosphene is arched dorsally. The small paracotylar foramina occur on either side of the circular cotylar rim.
REMARKS
The gracile structure of vertebrae with cl/naw ratio> 1, the presence of paracotylar foramina, as well as well-developed neural spine, the presence of prezygapophyseal processes, and the haemal keel developed in trunk vertebrae enable assignment to “ Colubrinae ”. The middle trunk vertebrae resemble those of the genus “ Coluber ” on the basis of the following combination of characters: 1) strongly craniocaudally elongated centrum of trunk vertebrae; 2) the vaulted neural arch; 3) the well-developed and narrow haemal keel; and 4) the prezygapophyseal processes which were probably long, based on the well-developed prezygapophyseal processes in anterior trunk vertebrae. “Colubrines” referred to the genus “ Coluber ” have frequently been reported from the European Neogene (e.g., Szyndlar 1991a, 2005, 2009, 2012; Szyndlar & Schleich 1993; Ivanov 2002a, b; Ivanov & Böhme 2011; Rage & Bailon 2005; Venczel 1994, 1998, 2001). There are four large “colubrine” species in the European Miocene: Coluber dolnicensis Szyndlar, 1987 (MN 3a-MN 4), C. caspioides Szyndlar & Schleich, 1993 (MN 3a-?MN 6), C. suevicus (Fraas, 1870) (MN 3a-MN 7+8) and C. pouchetii (de Rochebrune, 1880) (MN 4-MN 9). Coluber (s.l.) sp. differs from C. dolnicensis in the absence of a prominent step in the anterior part of the haemal keel ( Szyndlar 1987; Ivanov 2002a). It differs from C. caspioides in the smaller dimensions and the wider zygosphene ( Szyndlar & Schleich 1993; Ivanov 2002a). Coluber (s.l.) sp. differs from C. suevicus in the more vaulted neural arch and clearly smaller diameter of the prezygapophyseal articular facets ( Szyndlar & Böhme 1993; Ivanov 2002a). It differs from C. pouchetii in the apparently much lower neural spine and a cervical hypapophysis inclined posteroventrally rather than ventrally ( Augé & Rage 2000; Szyndlar 2009). Coluber (s.l.) sp., type 1 resembles C. hungaricus ( Bolkay, 1913) reported from the early middle Miocene (MN 6) of Germany ( Ivanov & Böhme 2011), the middle Miocene to early Pliocene (?MN 6-MN 9; MN 13-MN 14) of Hungary ( Venczel 1994, 1998, 2001), and the middle Miocene of Kazakhstan ( Ivanov et al. 2019) in: 1) the same dimensions; 2) most probably in the same height of the neural spine; 3) a similarly wide medial lobe of the zygosphene; and 4) a parapophysis of the same length as the diapophysis ( Venczel 1994; Szyndlar 2005). C. hungaricus displays a high intraspecific variability and the largest specimens have a dorsally thickened neural spine, parapophyses larger than diapophyses, as well as well-developed subcotylar tubercles ( Venczel 1998). Despite this variability, Coluber (s.l.) sp. cannot be attributed to C. hungaricus because of the presence of much more distinct, sharp, and straight subcentral ridges, a deeper notch in the caudal margin of the neural arch, as well as the smaller condyle. Because of the incomplete preservation of the rather scarce material we avoid a species level identification.
| MN |
Museu Nacional, Universidade Federal do Rio de Janeiro |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Scincoidea |
|
Family |
|
|
Genus |
Coluber
| Ivanov, Martin, Čerňanský, Andrej, Bonilla-Salomón, Isaac & Luján, Àngel Hernández 2020 |
Coluber
| IVANOV M. & MUSIL R. & BRZOBOHATY R. 2006: 229 |
Coluber
| IVANOV M. & MUSIL R. 2004: 229 |