
Gigantea urubambensis, Negrete, Lisandro H. L., Brusa, Francisco & Carbayo, Fernando, 2010
|
publication ID |
https://doi.org/10.5281/zenodo.194512 |
|
DOI |
https://doi.org/10.5281/zenodo.6198354 |
|
persistent identifier |
https://treatment.plazi.org/id/03C8A55E-FFB7-3562-7DF1-223BFEC2F8EA |
|
treatment provided by |
Plazi |
|
scientific name |
Gigantea urubambensis |
| status |
sp. nov. |
Gigantea urubambensis View in CoL sp. nov.
Material. Holotype, MLP 6083. Anterior region: transverse sections on 49 slides; horizontal sections on 18 slides; sagittal sections on 72 slides. Pre-pharyngeal region: transversal sections on 45 slides. Pharynx: sagittal sections on 81 slides. Copulatory apparatus: sagittal sections on 99 slides. Rest of body in 70%–100% ethanol.
Diagnosis. Body large (length: 160 mm, maximum width: 13 mm, maximum height: 5.5 mm), elongated and lanceolated; dorsally with two thin paramedian reddish stripes along a median yellowish orange band, black lateral bands; body margins whitish; eyes initially marginal and uniserial encircling the anterior tip, backwards becoming dorsal and pluriserial with clear halos along the black lateral bands and yellowish orange median band; dorsal epithelium ciliated in the cephalic region; glandular margin absent; CMI, 5.0– 6.9%; pharynx collar-type; esophagus absent; testes very dorsal, between dorsal diagonal and supra-intestinal parenchymatic muscle layers; prostatic vesicle absent; penis papilla large, ventrally projected from the roof of the male atrium, acentrally traversed by the ejaculatory duct; glandular ridges on penis papilla absent; ovovitelline ducts ascend posterior to the female atrium; common glandular ovovitelline duct absent; female atrium wide.
Type locality. Tropical rainforest near Camisea (11º 51’ S; 72º 56’ W) in the Lower Urubamba Region, in Cusco, Peru.
Etymology. The specific name refers to the Lower Urubamba River Valley of southern Peru, where the specimen was collected.
Description. External features. The live specimen was 160 mm long and 13 mm in maximum width at pharynx level. The body is elongated and lanceolated, with a blunt rear end. After fixation, the worm was 120 mm long and 12 mm in maximum width (10% width:length ratio). Maximum height was 5.5 mm, 63 mm from the anterior end (53.0% of body length). Mouth and gonopore are located at 70 mm (58.3%) and 90 mm (75.0%), respectively, from the anterior end.
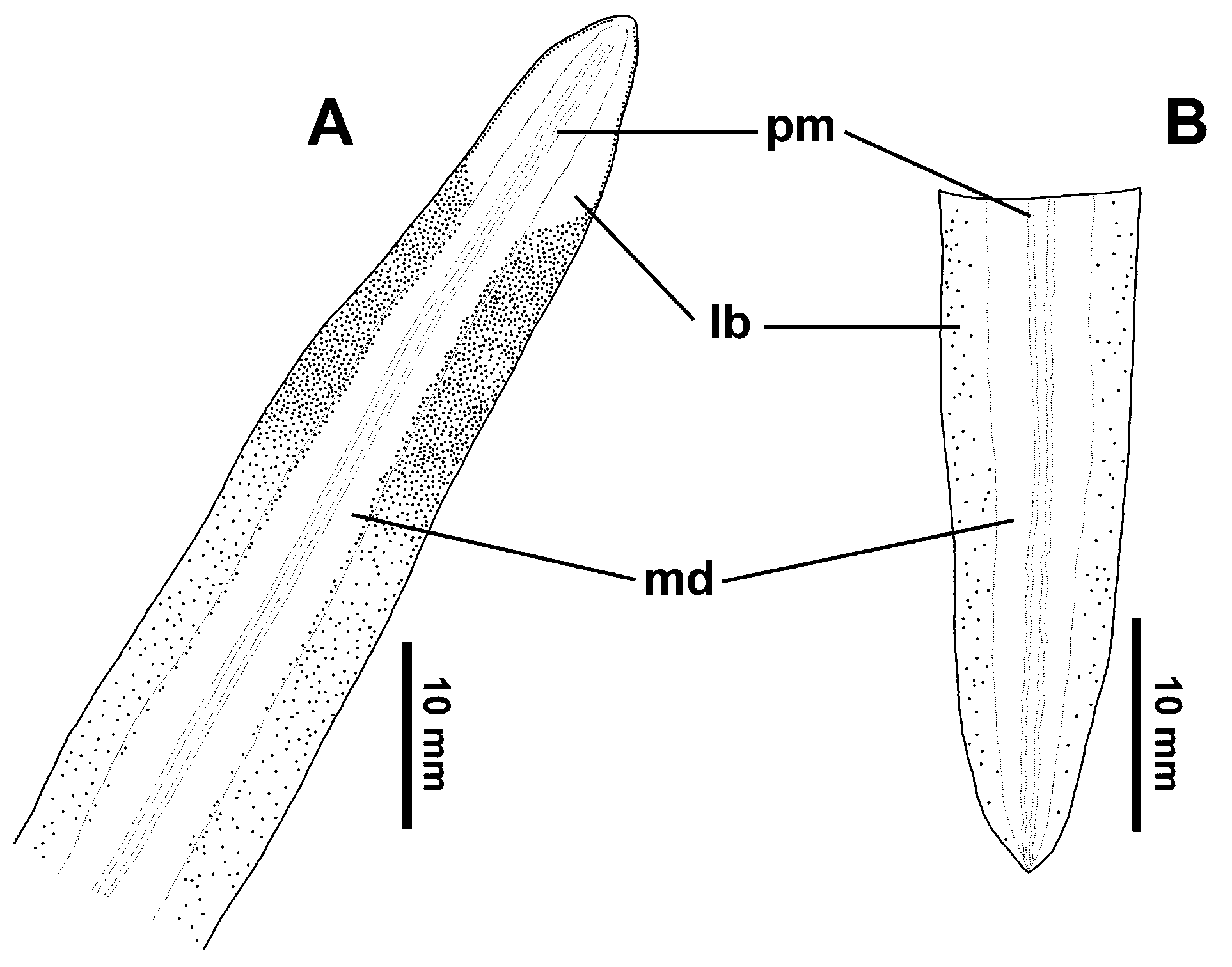
The dorsal side shows two thin paramedian reddish stripes along a median yellowish orange band (41% of body width); two black lateral bands (42% of body width). Further markings consist of two marginal whitish stripes (17% of body width) ( Fig. 1 View FIGURE 1 ). The ventral side is light whitish gray.
Eyes initially marginal and uniserial, encircling the anterior tip. At 9 mm from the anterior tip (8% of body length) they become dorsal and pluriserial with clear halos along the black lateral bands and the margins of the median yellowish orange band ( Fig. 2 View FIGURE 2 A). Posterior to 48 mm from the anterior tip (40% of body length), the eyes become scattered along the black lateral bands to reach the posterior body region ( Fig. 2 View FIGURE 2 B).
Epidermis and secretions. The ventral epithelium is ciliated across the entire width of the body, thus forming a broad creeping sole. The dorsal epithelium is only ciliated in the anterior region. At the level of the pre-pharyngeal region, the epithelium is 41 µm high on the dorsal side, 36 µm ventrally and 38 µm at the body margins. Erythrophilic granular glands open through the entire epithelium but a glandular margin is absent. Abundant rhabditogenic cells traverse the entire epithelium, being much more abundant on the dorsal side than on the ventral. In the cephalic region, cianophilic granular secretions traverse the latero-ventral epithelium.
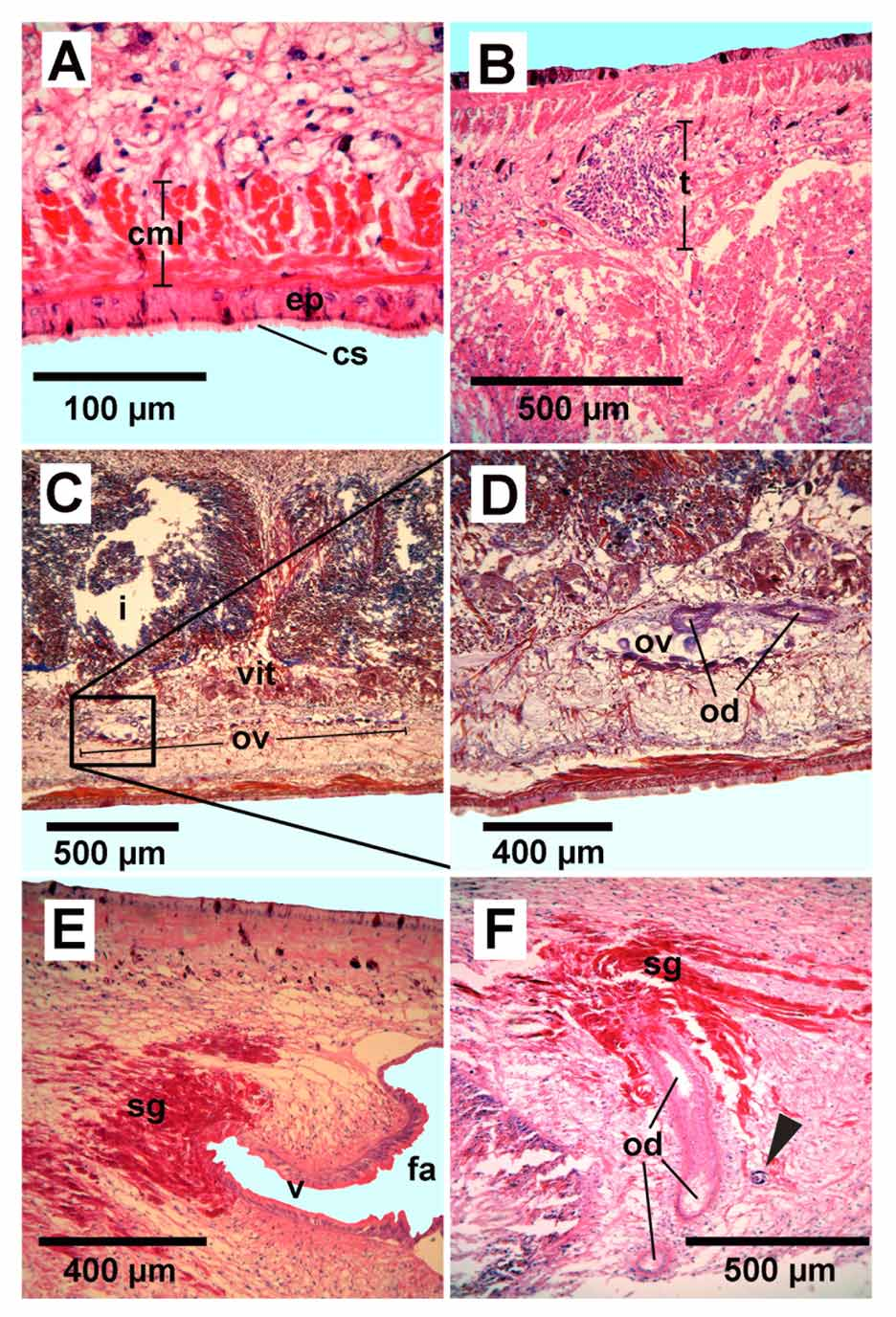
Cutaneous musculature. The cutaneous musculature is formed by the three layers that are typical of the Geoplaninae : an external circular layer, a double diagonal layer with decussated bundles, and a longitudinal layer also organized in bundles ( Figs. 3 View FIGURE 3 A, 4A). In the pre-pharyngeal region, the circular layer is thicker dorsally than ventrally, being 34 µm and 15 µm respectively. The diagonal musculature is equally thick at the dorsal and ventral regions, ranging from 19 to 34 µm. The longitudinal muscular layer is 53 to 97 µm thick, equally developed dorsally and ventrally. CMI ranges between 5.0% and 6.9%. The cutaneous muscle layers grow thinner towards the sides of the body, with the longitudinal muscle being most conspicuous. In the cephalic region the layers also grow thinner with no changes in orientation of the muscle fibers, but the anterior most 0.5 mm was missing.
Parenchymal musculature. In the pre-pharyngeal region, the parenchymal muscle fibers are arranged in various directions, with the dorso-ventral ones being most abundant. Other parenchymal fibers are arranged in three layers that together correspond to 10% of body height: (1) a dorsal diagonal layer with decussated fibers (22.5 µm thick) located under the dorsal submuscular peripheral nerve net; (2) a supra-intestinal transverse layer (125 µm thick); (3) a sub-intestinal transverse layer (100 µm thick) ( Fig. 3 View FIGURE 3 A). The dorsal diagonal layer is formed by bundles of 4–7 fibers each. The other two layers do not present bundles. Numerous oblique fibers run between the intestinal branches, from dorsal to ventral epidermis extending to the body margins. Muscular fibers become scarce towards the anterior end of the body and disappear near the tip, whose first 0.5 mm was missing.
Digestive system. The mouth is located in the middle of the pharyngeal pouch. The pharynx, collar type, is 7.6 mm long (~6% of body length) ( Fig. 3 View FIGURE 3 B) and strongly folded. An esophagus is absent. The lining epithelium of the pharyngeal pouch is flat, not ciliated, and columnar around the mouth; it is surrounded by sub-epithelial muscle layer, one-fiber thick, which is longitudinal at the pharyngeal region, and decussated posteriorly to it.
The outer pharyngeal epithelium is squamous, with the nuclei sunken below its muscularis, and columnar with basal nuclei at the pharyngeal insertion. It is traversed by two types of granular secretions, erythrophilic and cianophilic. The outer musculature comprises a longitudinal layer (10 µm thick) followed by a circular muscle (19–24 µm thick). The epithelium of the pharyngeal lumen is squamous, with the nuclei sunken into the parenchyma; a circular muscular layer (190–385 µm thick) lies underneath, with some interspersed longitudinal fibers.
Male reproductive system. The testes are rounded and located dorsally between the dorsal diagonal and the supra-intestinal parenchymatic muscle layers, occupying about one-sixth of the dorso-ventral height in the pre-pharyngeal region ( Figs. 3 View FIGURE 3 A, 4B). The follicles are arranged in 1–5 rows on either side of the body. The posterior-most testes are lateral to the pharynx. The anterior-most and the posterior-most testes are located at 10.0% and 59.6%, respectively, from the anterior tip.
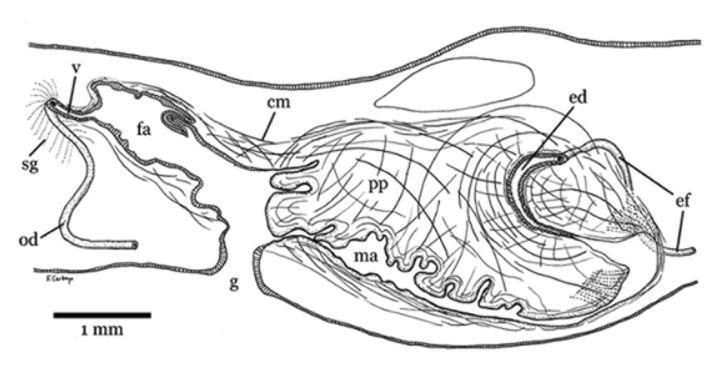
The efferent ducts run ventrally to the parenchymal subintestinal muscular layer, above the ovovitelline ducts and slightly internal to them ( Fig. 3 View FIGURE 3 A). Their epithelium is cuboidal and ciliated, and the distal lumen contains spermatozoa. They are surrounded by a decussated 13 µm thick muscle layer. In the vicinity of the proximal region of the male atrium, the efferent ducts turn dorsally, then curve to the sagittal plane, penetrate horizontally the common muscle coat, communicate with each other, and open into the ejaculatory duct ( Fig. 6 View FIGURE 6 ). The latter is a slightly dilated canal, C-shaped in lateral view, with its the distal section oriented forward ( Figs. 5 View FIGURE 5 , 6 View FIGURE 6 ). It traverses the penis papilla acentrally and opens near the ventral insertion of the latter. The columnar and ciliated epithelium of the ejaculatory duct is 18 µm high. The apical portion of its cells is inclined towards the opening of the duct. Scarce cells with an erythrophilic granular secretion open onto the lumen. The ejaculatory duct is surrounded by a one-fiber thick circular muscle layer, followed by a 10 µmthick layer of longitudinal fibers.
The penis papilla, large and distally folded, occupies the entire male atrium and projects ventrally from its roof. The epithelium of the penis papilla is columnar with basal nuclei. If cilia are present, then they are inconspicuous due to secretion in male atrium ( Fig. 5 View FIGURE 5 ). Around the opening of the ejaculatory duct the epithelium of the penis papilla is squamous, non ciliated, traversed by numerous cells with an erythrophilic granular secretion. The muscularis of the penis papilla comprises a sub-epithelial circular layer, 25 µm thick, followed by a longitudinal layer, 70 µm thick. The parenchyma of the penis papilla possesses abundant muscle fibers arranged in several directions.
The male atrium is a long cavity completely occupied by the penis papilla. It is twice as long as the female atrium. Its epithelium is squamous, apically erythrophilic. The presence of secretion in the atrial lumen prevented us from discerning whether this epithelium is ciliated or not ( Fig. 5 View FIGURE 5 ). The epithelium is underlain by a circular muscle layer, 12–17 µm thick, followed by a longitudinal layer, 63–70 µm thick. Numerous cells with erythrophilic granular secretion, underneath the longitudinal muscle layer, open together in the transition area between male and female atrium.
Female reproductive system. The ventral, elongated ovaries (1.2 mm x 0.15 mm) are located between the sub-intestinal muscle layer and ventral nerve plate ( Fig. 4 View FIGURE 4 C, D), at a distance equal to 8% of body length from the anterior end of the body. The vitellaria are well developed, distributed among and beneath the intestinal branches ( Figs. 3 View FIGURE 3 A, 4C).
The ovovitelline ducts arise from the antero-dorsal part of the ovaries ( Fig. 4 View FIGURE 4 C, D). The ducts run within the parenchymatic sub-intestinal muscle layer. They run posterior to the gonopore and to the female atrium, and then turn dorsally to join directly with the vagina ( Fig. 6 View FIGURE 6 ). A common glandular ovovitelline duct is absent. The vagina, formed by an almost horizontal and elongated diverticulum of the female atrium, opens in the proximal region of the latter. The female atrium is an oblique and wide cavity, 1.3 mm long occupying at least the dorsal half of the body height ( Figs. 5 View FIGURE 5 , 6 View FIGURE 6 ), presenting some folds that project into its lumen.
The ovovitelline ducts have a ciliated epithelium and the distal third of the ascendant portion receives the shell glands ( Fig. 4 View FIGURE 4 F). The vagina has a non-ciliated, apically erythrophilic, pseudo-stratified epithelium, with the distal portion of its cells inclined towards the ovoviteline ducts ( Fig. 4 View FIGURE 4 E). This epithelium overlies a decussated muscular layer, 8 µm thick. The lining epithelium of the female atrium is pseudo-stratified, nonciliated, apically erythrophilic. Beneath the epithelium there is a thin decussated muscle layer, followed by a circular layer and another with intermingled longitudinal and circular muscle fibers. The proximal ventral epithelium of the female atrium is traversed by cells with an erythrophilic granular secretion, and their bodies are beneath the intermingled muscle fibers. The gonopore canal is straight and lined with a columnar, ciliated epithelium.
The common muscle coat is formed by a thick layer of longitudinal and oblique fibers, more developed in the male organ than the female.
Parasitism. Nematode larvae were found in the parenchyma, both near the ejaculatory duct and below the vagina ( Fig. 4 View FIGURE 4 F).



| MLP |
Museo de La Plata |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Geoplaninae |
|
Genus |