
Boiga beddomei ( Wall, 1909 )
|
publication ID |
https://doi.org/10.11646/zootaxa.4779.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:0B6F641C-424E-4042-A9B6-A130C58935AB |
|
DOI |
https://doi.org/10.5281/zenodo.3851852 |
|
persistent identifier |
https://treatment.plazi.org/id/03CB8788-EB20-FFF8-FF04-7EB3FDE9396C |
|
treatment provided by |
Plazi |
|
scientific name |
Boiga beddomei ( Wall, 1909 ) |
| status |
|
Boiga beddomei ( Wall, 1909) View in CoL
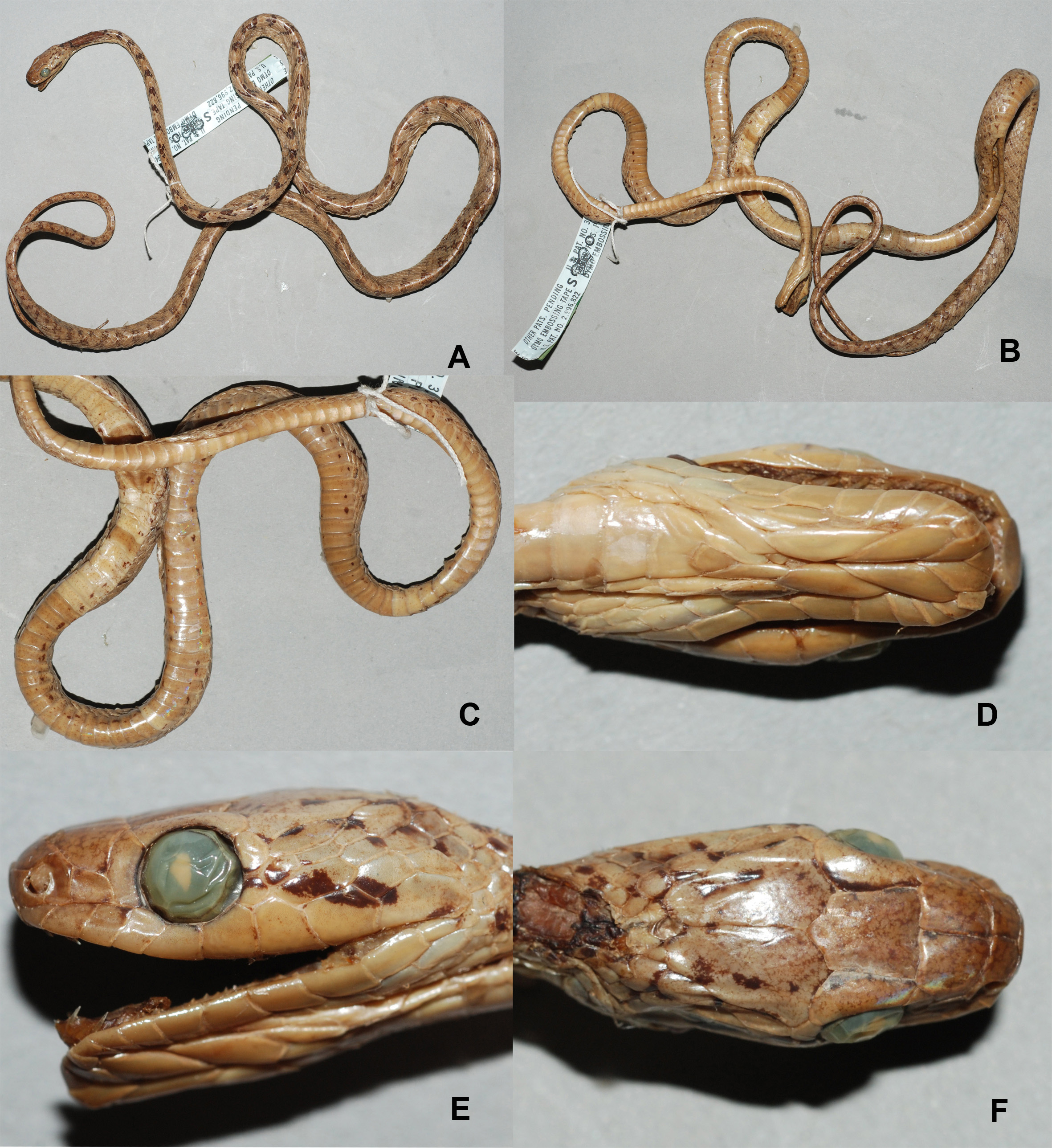
( Tables 3–4; Fig. 4 View FIGURE 4 , 9C View FIGURE 9 )
Dipsadomorphus beddomei Wall, 1909
Dipsadomorphus beddomei — Wall, 1919
Boiga beddomei— Whitaker & Captain, 2004 part
Specimens examined. Females (n=4). Lectotype (designated herein): India. Maharashtra. BMNH 69.8 .28.123, from “Matheran”; collected by Dr. Leith, probably Andrew Henderson Leith ( 1848-1919).
Other specimens: BNHS 1854 and 1854 Matheran; BNHS 1896 Khandala.
Taxonomic History. Boiga beddomei was described by Wall (1909) as Dipsadomorphus beddomei during his revision of the B. ceylonensis group. The description was based on a series of specimens from India and Sri Lanka ( Vogel & Ganesh 2013; Wallach et al. 2014). Annandale (1909) and Smith (1943) synonymized it with B. ceylonensis . However later authors (Taylor, 1950) considered it to be valid. Manamendra-Arachchi et al. (2007) was misled that its types were lost and synonymised Boiga ranawanei (see Samarawickrama et al. 2005) with it ( Vogel & Ganesh 2013).
As the type series of Dipsadomorphus beddomei Wall, 1909 is a mixture of several populations ( Wall, 1909) and as it is part of a species complex ( Vogel & Ganesh, 2013), it is necessary to designate a lectotype to clarify the taxonomic status. Thus, as per Art. 74.7 of the Code (ICZN, 1999), we hereby designate an adult specimen BMNH 69.8.28.123 from a precisely named Indian locality “Matheran” as the lectotype of Dipsadomorphus beddomei . The chosen lectotype, is the only specimen of the type series originating from India.
Etymology. Patronym named in genitive singular case honouring Col. Richard Henry Beddome ( 1830–1911), one of the most prominent herpetologists of South Asia.
Diagnosis (redefined herein). A species of Boiga inhabiting the Western Ghats, defined by the following combination of characters: 19 midbody scale rows (vs. 21–23 in B. nuchalis , B. dightoni , B. andamanensis ); predominantly yellowish-brown dorsum (vs. greenish in B. flaviviridis ; variable in B. andamanensis ); vertebrals strongly enlarged (vs. mildly enlarged in B. barnesii ); crown unpatterned (vs. with blackish-brown blotches in B. thackerayi , brownish blotches in B. ceylonensis ); subcaudals> 110 (vs. < 110 in B. ceylonensis , B. thackerayi ); venter densely speckled with dark brown (vs. uniform in B. flaviviridis ); ventrolateral region devoid of large white and black blotches (vs. present in B. thackerayi , B. barnesii ), but has a series of spots on both tips of each ventral scale (vs. venter unpatterned in B. andamanensis , B. flaviviridis ).
Description of the lectotype. A medium-sized snake reaching 809 mm total length, snout-vent length: 624 mm, tail length: 185 mm, long tail (relative tail length 22.9%); slender habitus, thin neck, wide head; 19–19–15 scale rows; rostral visible from above; preocular 1, subequal to loreal; postoculars 2; loreal 1; supralabials 8/9, with 3 rd– 5 th ones touching eye; infralabials 11/12, with 1–5 touching chin shields; temporals 10/8; preventral 1; ventrals 247, angulate laterally; cloacal 1; subcaudals 117 pairs. Dorsal colour brownish grey, with 109 (81 on body, 28 on tail) blackish crossbars; crossbars covering 2–3 scales in size, extending either sides up to 2–3 scale rows across; interspaces often with sparse dark dots; crown without any markings on top; a thin postocular stripe up to the jaw angle; labials, chin and venter creamy white, finely spotted with darker hue; venter off-white, with a consequent series of brown spots resembling dotted lines, along the ventrolateral edges on both the sides.
Variation. Other specimens agreeing with the lectotype in most respects and showing the following intraspecific variations: snout-vent length 756–822 mm; tail length 187–219 (incomplete); (18)19–19–15 (14) scale rows; ventrals 238–252; subcaudals 84–87+? (incomplete); temporals 10 in all specimens on both sides; supralabials 8–9, rarely with 4 th– 6 th touching eye; infralabials 10–11; dorsal bands on body 76–81, on tail 31–33.
Distribution and natural history. This species occurs only in the hills of northern Western Ghats, north of Goa Gap up to Surat Dangs, where it inhabits deciduous and semi-evergreen forests (Chikane & Bosle 2012; Whitaker & Captain 2004 part) (Fig. 13). The status of the population from Sri Lanka (see Taylor 1950; Das & De Silva 2005; Samarawickrama et al. 2005; Somaweera 2006; Manamendra-Arachchi & Pethiyagoda 2007) warrants further studies (see below).
Comments. Manamendra-Arachchi & Pethiyagoda (2007) considered B. ranawanei to be synonymous with B. beddomei as they conceived this species. They created an objective synonymy between these two taxa by designating the holotype of B. ranawanei as the neotype of B. beddomei . As we located without ambiguity the original type series of B. beddomei (see also Vogel & Ganesh 2013), the neotype designation done by Manamendra-Arachchi & Pethiyagoda (2007) becomes automatically invalid ( fide, Art. 75.8 ICZN 1999). The type series of B. beddomei is a mixture of two populations, both from India and Sri Lanka. We have here designated the only Indian syntype as the lectotype of B. beddomei . The status of the remaining, Sri Lankan specimens, i.e. the paralectotypes of ‘ B. beddomei ’ sensu lato in BMNH holdings (BMNH 46.12.2 Ceylon, BMNH 93.10.6.3_4, BMNH 93.10.6.3 and BMNH 93.10.6.4, all from “Trincomalu” (now Trincomalee or Tirukonamalai, in Eastern Province, Sri Lanka.) as well as the nominal Sri Lankan species B. rananwanei await further taxonomic works, pending better resolution of this group and is deferred to future works.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.