
Symphysanodon pitondelafournaisei
|
publication ID |
https://doi.org/10.5281/zenodo.278702 |
|
DOI |
https://doi.org/10.5281/zenodo.5696920 |
|
persistent identifier |
https://treatment.plazi.org/id/03CCCD04-9C51-FFF6-4DB7-963FCCF2FA3B |
|
treatment provided by |
Plazi |
|
scientific name |
Symphysanodon pitondelafournaisei |
| status |
|
Symphysanodon pitondelafournaisei View in CoL
It seems appropriate to comment here on another recently described species of Symphysanodon , S. pitondelafournaisei Quéro, Spitz, and Vayne, 2009 , known from two specimens (90 & 99 mm SL) collected at the surface off Réunion Island (Mascarene Islands), southwestern Indian Ocean (at 21°07’S, 55°32’E), in April 2007, after an eruption of the volcano Piton de la Fournaise. Although we have not examined either specimen, radiographs of the holotype of S. pitondelafournaisei (MNHN 2008-0151, 99 mm SL) show it to have 25 (10 precaudal + 15 caudal) vertebrae, parapophyses on the first caudal vertebra, configuration of supraneural bones, anterior neural spines, and anterior dorsal pterygiophores 0/0/0 + 2 + 1/1/1/, three epural bones, and 17 (9 + 8) principal caudal-fin rays (in contrast with the count of 19 given in the original description).
Symphysanodon pitondelafournaisei is distinguishable from all other known species of Symphysanodon by the following combination of characters: parapophyses present on the first caudal vertebra, pectoral-fin rays 15, total number of gillrakers on first arch 34 to 36, tubed lateral-line scales 48 to 50, sum of total number of gillrakers plus tubed lateral-line scales 84. Based on the color photograph published in the original description ( Quéro et al., 2009:75, fig. 2), S. pitondelafournaisei displays a pattern of coloration which distinguishes it from S. berryi , S. disii , S. katayamai , S. maunaloae , S. typus , and S. xanthopterygion , and which may allow it to be separated from the other species of Symphysanodon for which no information on live coloration is currently available.
In the original description ( Quéro et al., 2009:74) the coloration was described as:
D’après photographie à l’état frais... le corps de l’holotype est saumon dorsalement, argenté ventralement; les nageoires dorsale, anale et caudale sont jaunes. L’iris est jaunâtre. Une tache jaune au-dessus du bord antérieur de la base des pectorales semble être présente; le corps du paratype et ses nageoires sont brun-rougeâtre. Dans l’alcool le corps de l’holotype est gris–jaunâtre, plus foncé dorsalement, plus argenté ventralement; le paratype a le dos gris-saumoné.
(= According to the photograph of the fresh state... the body of the holotype is salmon dorsally, silvery ventrally; dorsal, anal and caudal fins are yellow. The iris is yellowish. A yellow spot above the anterior border of the base of the pectoral [fin] apparently present; the body of the paratype and its fins are reddish brown. In alcohol the body of the holotype is yellowish grey, darker dorsally, more silvery ventrally; the paratype has the back grey salmon.)
The published color photograph ( Quéro et al., 2009: 75, fig. 2) shows the head and body of S. pitondelafournaisei to be mainly reddish with a broad yellow band running from posterior part of opercle to caudal peduncle where it narrows appreciably and becomes indistinct, iris mainly yellow but with red dorsally, dorsal fin yellow green, pectoral fin red orange, anal and pelvic fins pallid to transparent, both lobes of caudal fin mainly bright yellow basally, yellow green distally with produced filamentous ends of lobes reddish.
Sexual dimorphism in Symphysanodon . As noted by Anderson (1970), S. berryi is sexually dimorphic in dentition and in lengths of the pelvic fin and lobes of the caudal fin. Length of pelvic fin varying in females from 21 to 25 % SL and in males more than ca. 85 mm SL from 30 to>87 % SL. The caudal fin is deeply forked with upper and lower lobes produced into long filaments in large males, increasing in length with increase in SL. Length of upper lobe of caudal fin varying in females more than ca. 80 mm SL from 30 to>35 % SL and in males more than ca. 85 mm SL from 34 to>128% SL. Length of lower lobe of caudal fin varying in females more than ca. 80 mm SL from 30 to>34% SL and in males more than ca. 85 mm SL from 32 to>111 % SL. In addition, the dentition is better developed and coarser in males than in females.
Anderson (1970:338) found that seven ( 62–86 mm SL) of the 30 specimens of S. maunaloae that he examined “have the pelvic fin well produced (with the medial branch of the first pelvic soft ray reaching anterior part of base of anal fin or beyond), whereas in the other specimens ( 47–86 mm SL) the pelvic fin, although usually produced, falls short of the anal fin.” Although he had only limited success at determining the sex of the available specimens, he wrote that: “It seems, however, that the well-produced pelvic fin is a male characteristic (as it is in S. berryi ).” Tameka (1982:372) noted that: “First soft ray of pelvic fin elongated in male, reaching backward beyond base of anal fin or to caudal fin. Pelvic fin of female not elongated.” Anderson (1970) reported that the pelvic fin varied from ca. 22 to>54 % SL. In 16 ( 79–141 mm SL) of an additional 48 specimens, Anderson and Springer (2005) found the pelvic fin to be considerably produced, ranging from ca. 42 to>80 % SL. Gross examination of the gonads of two of those specimens (120 & 128 mm SL) showed them to be males.
Anderson (1970) reported the length of upper lobe of caudal fin of S. maunaloae to range from ca. 31.2 to ca. 48.9 % SL and the length of lower lobe to range from ca. 30.0 to ca. 43.1 % SL. Measurements of the additional specimens mentioned by Anderson and Springer (2005) extend those ranges to 29 to ca. 75 % SL for upper lobe and ca. 28 to ca. 76 % SL for lower lobe. Two of those specimens (120 & 128 mm SL) with very long caudal-fin lobes are males (see above). Well-produced pelvic fins and caudal-fin lobes appear to be characteristic of males in S. maunaloae , as they are in S. berryi .
Based on histological examination two (134 & 136 mm SL) of the six known specimens of S. rhax are female ( Anderson and Springer, 2005). Those two and two others have short pelvic fins (20->26% SL); the remaining two (136 & 144 mm SL) have produced pelvic fins (>64 &> 67 % SL). “Because both specimens known to be female by histological examination have short pelvic fins and because females of the apparent closest relative of this species, S. berryi , have short pelvic fins (in contrast with males which frequently have very well-produced pelvic fins), it is reasonable to assume that the individuals of S. rhax with produced pelvic fins are males” ( Anderson and Springer, 2005:21). As noted above, S. berryi also exhibits sexual dimorphism in lengths of caudal-fin lobes. Damage to both caudal-fin lobes on all six specimens of S. rhax precludes answering the question of sexual dimorphism in this character.
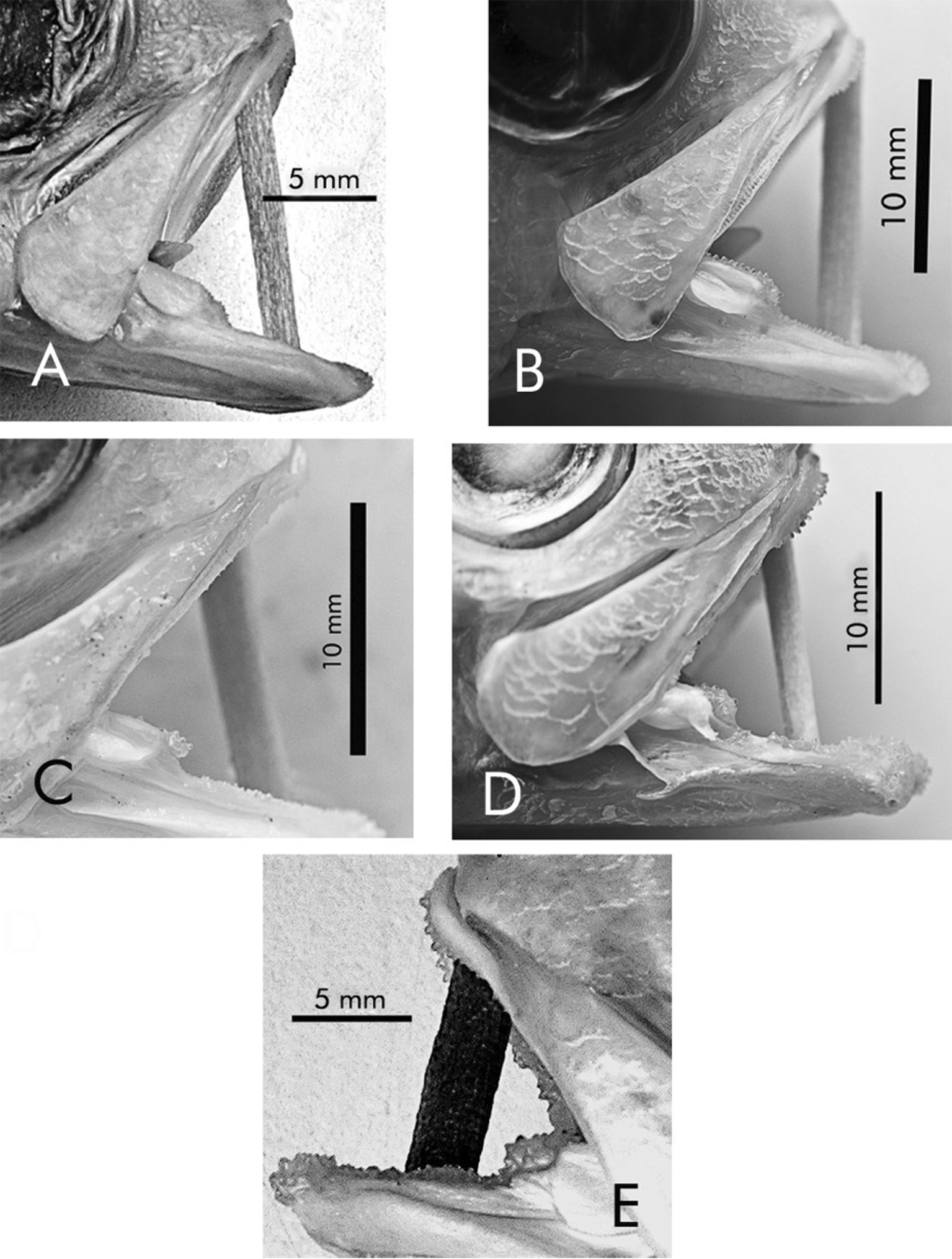
Sex was determined histologically in all five specimens of S. disii (one female, 146 mm SL; four males, 148– 163 mm SL). The dentition is much better developed in the males than in the female (see above for d e tails of dentition in S. disii , and Fig. 2 View FIGURE 2 ), suggesting that there are differences in diets or dietary preferences between the sexes or that these distinctions in dentition may relate to spawning behavior. The sexual dimorphism displayed in the pelvic and caudal fins of berryi , maunaloae , and rhax may function in sexual recognition or may indicate that males and females exploit, at least in part, somewhat different habitats or under certain conditions display different patterns of behavior. We are aware of no other species of Symphysanodon that have obvious sexual dimorphism. This is not surprising in view of the fact that most species for which sexual dimorphism has not been observed are represented in museum collections by a small or relatively small numbers of juvenile and adult specimens.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |