
Austrodalyellia ariena
|
publication ID |
https://doi.org/10.5281/zenodo.156078 |
|
DOI |
https://doi.org/10.5281/zenodo.6279081 |
|
persistent identifier |
https://treatment.plazi.org/id/03CE878B-FF8A-5D2C-E20F-FCDD20127AE5 |
|
treatment provided by |
Plazi |
|
scientific name |
Austrodalyellia ariena |
| status |
|
Austrodalyellia ariena View in CoL . sp. nov. ( Figs. 16 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 )
Material Examined. Approximately twentyone specimens studied alive, four made into wholemounts, and eight sectioned.
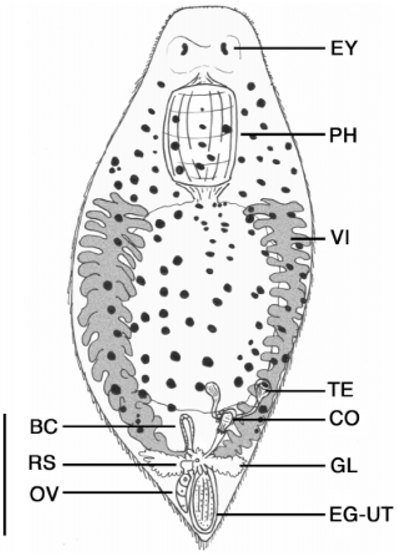
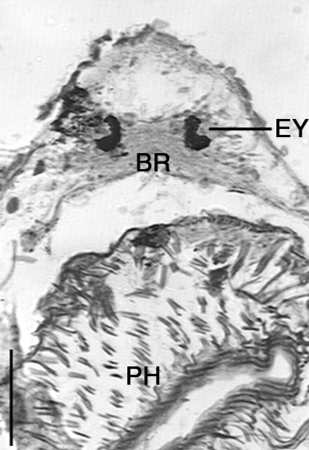
Description. Animals are up to 1.6 mm long and 700 µm wide with a rounded head, wide body and rounded to pointed caudal region ( Fig. 1 View FIGURE 1 ). Small tail sometimes present. Body often transparent except for glandular intestine and numerous red to brown pigment spots covering body. The epidermis is a ciliated epithelium, with cuboidal cells to 8 um high in section over most of the animal and containing 35 basophilic rhabdites per cell. Longitudinal, circular and possible diagonal muscle fibres present below epidermis. Anteriorly, paired kidneyshaped eyes (19 µm long) of numerous black pigment spots cover a bilobed brain ( Fig. 2 View FIGURE 2 ). Posterior to the eyes is a large, muscular doliiform pharynx, usually to onefourth body length, leading posteriorly into a short eosophagus and finally into a saccate intestine. Gut often contains algal cells and annelid setae. Branching protonephridia are lateral to the pharynx and intestine, extending from head to tail.
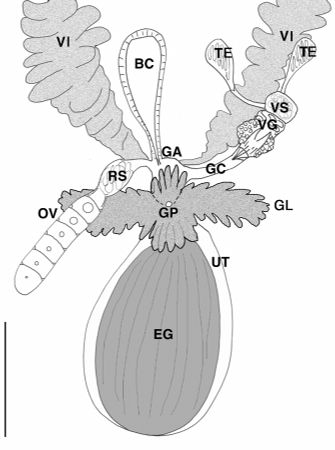
The female reproductive system consists of paired vitellaria, solitary ovary, receptaculum seminis, uterus, bursa copulatrix, genital atrium and common gonopore ( Figs. 1 View FIGURE 1 , 34 View FIGURE 3 View FIGURE 4 ). Paired vitellaria present as two longitudinal bands beginning lateral to esophagus and extending to caudal end. Vitellaria consist of numerous fingerlike processes (to 96 µm long) and often extend dorsally over intestine. Caudally, short vitelline ducts connect to a common genital atrium. Extending anteriorly from the genital atrium is a solitary, elongate ovary often located on the left side of the body. Distally the ovary leads into a short oviduct before expanding into an oval receptaculum seminis often containing sperm. A small, muscular bursa copulatrix (55 µm long) with a short distal tube, also muscular, leading to the genital atrium was located at the body midline. In several specimens the bursa was obscured by parenchymal pigments. The region around the genital atrium is highly glandular with numerous digitiform glands (stains intensely with aniline blue) that often conceal the common gonopore and all ducts leading to it. Posterior to the genital pore is a thinwalled uterus containing an egg up to 200 µm long. Eggs are bright orange and highly sculptured. In one specimen (egg size 192 µm long and 128 µm wide) the dorsal sculpturing (observed from the dorsal side of the animal) consisted of 15 raised rows of numerous small spheres 1.6 µm to 2.4 µm in diameter. Raised rows were oriented longitudinally, separated by depressed areas 78 µm wide and often filled with numerous small spheres of variable size. Egg shell thickness was 1.8 um in depressed areas and up to 9 um in the raised ridges.
The male reproductive system consists of paired testes, vasa deferentia, vesicula granulorum, vesicula seminalis, a sclerotic copulatory organ, genital atrium and common gonopore ( Figs. 1 View FIGURE 1 , 35 View FIGURE 3 View FIGURE 4 View FIGURE 5 ). The paired testes are ventral to the vitellaria and located on the posterolateral border of the intestine on the right side of the body. Each testis is round and compact and gives rise to a vas deferens that leads to the copulatory organ. The organ contains an anterior saclike vesicula seminalis atop a compact glandular vesicula granulorum. The base of the organ was ensheathed in its own longitudinal musculature while some fibres originating from the bodywall also appeared to supply the organ. A sclerotic copulatory apparatus was present at the distal end of the organ.
When relaxed, the copulatory apparatus is housed within a muscular bulb that leads into a thinwalled genital canal ( Figs. 3 View FIGURE 3 , 5 View FIGURE 5 A). Several spinelike structures could be seen protruding from the distal end of the canal. In heavily squashed whole mounts, the entire structure is bilaterally symmetric and consists of three layers of complex tines: a single dorsal tine, paired median tines, and paired ventral tines ( Figs. 5 View FIGURE 5 , 6 View FIGURE 6 ). The dorsal tine, ca. 59 µm long, is in the shape of a “V” and constructed of thin bars. A thin membrane exists between the bars for the length of the tine except around 50 % length where an aperture (8 µm by 5 µm) is present. Proximally, the left and right base of the dorsal tine is connected to the respective left and right tines immediately ventral to it. The paired median tines are lanceolateshaped structures, ca. 58 µm long, constructed similar to the dorsal tine but with a larger and more triangularshaped aperture (18 µm long) at ca. 60% length. The paired ventral tines, ca. 58 µm long, are somewhat bladeshaped with two apertures: a lateral circular aperture to 7 um diameter and a medial elliptic aperture to 4 µm wide. A small curved bar extends from the most medial side of the circular aperture to the lateral side of the tine. The tip and base of the bladeshaped tine are perforated with tiny holes.
When extended, the copulatory organ appears highly flexible. Tines can twist and bend longitudinally. In several specimens, the dorsal tine and paired ventral tines were bent outward at approximately 25% tine length (from the base) and had peeled away from the paired medial tines. In other specimens, the dorsal tine remained straight and the median and ventral tines had peeled away.
Remarks. Austrodalyellia ariena gen. nov. sp. nov. conforms to all general morphological characters of the related freshwater genera Dalyellia , Gieysztoria and Microdalyellia : large anterior doliiform pharynx, saccate gut, paired testes, sclerotic male copulatory organ, paired vitellaria, solitary ovary, bursa copulatrix, seminal vesicle, and uterus. The topology of the male reproductive system is perhaps more reminiscent of species of Gieysztoria , with posterior testes and short vas deferentia leading to a compact vesicula seminalis and vesicula granulorum. The cuticular apparatus, however, bears little resemblance to the stylets of other dalyellioids.
General stylet morphology in Dalyellia and Microdalyellia is covered extensively in Luther (1955) and consists of several tines: two proximal handles (Stiel) connected by a cross piece (Querbalken), a dorsal and ventral median piece (Rinne), and two lateral distal branches (Endäste) with spines. Similarly, the stylet of the marine dalyelliid, Jensenia angulata (Jensen 1878) , consists of the same basic structures minus the ventral median piece ( Ehlers 1990). In Austrodalyellia , the copulatory stylet also consists of several tines in relatively similar positions, though their morphology differs considerably from that of other species. The stylet is bilaterally symmetric and constructed of five tines: two layers of ventral tines on either side of the midline of a single dorsal tine. The dorsal tine is connected to all four pieces at its paired proximal base.
Homology of individual tines with those of other species is difficult to assess. Based on relative shape and position, the dorsal tine may be homologous with the dorsal median piece of species of Dalyellia , Microdalyellia , Jensenia and perhaps some species of Gieysztoria (see Rogozin 1995). The dorsal centerpiece is characteristically a solid, distal triangular expansion of the proximal handles. The dorsal tine of Austrodalyellia is also triangular and relatively solid (the thin membrane often appears absent under strong light) with the exception of the medial aperture. The paired short proximal ends of the stylet are also conceivably derived from the elongate handles characteristic of species of Dalyellia and Microdalyellia among others. The homology of the median and ventral distal tines is more difficult to assess. Their structure is similar to the distal tine but their shape is unlike any of the stylet branches present in other species.
Functionally, the stylet of Austrodalyellia appears highly flexible, and while all individual tines can bend along their length, it is the dorsal and ventral tines that often ‘peel’ away from medial tines during stylet discharge (during fixation; Figs. 5 View FIGURE 5 B, 6B). The effect is reminiscent of the opening of the stylet of Castrella groenlandica Riedel, 1932 (see Fig. N1,p. 275, Luther 1955). Movements of individual tines other than lateral bending appears restricted presumably because all tines are branches of the common base and devoid of joints. Unfortunately, muscle attachments to individual tines could not be ascertained from histological sections, so their range of possible movements is unknown. However, the perforated appearance of the proximal handles and ventral tines might indicate insertion points for retractor muscles.
There appears to be minimal variation in the structure or position of the copulatory stylet among the examined specimens. However, it was difficult to make good squash mounts of the stylet, and unfortunately, in the majority of specimens examined, the stylet was ‘peeled', making observations of individual tines extremely difficult. Attempts to isolate the stylet by dissolving away soft tissue using dilute sodium hypochlorite also dissolved much of the stylet, especially the thin membraneous sheet that connects the tine bars. Future attempts to isolate and observe the stylet using scanning electron microscopy might provide more useful taxonomic characters than can be ascertained with light optics.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |