
Sesieutes Simon, 1897
|
publication ID |
https://doi.org/ 10.1080/00222933.2012.742165 |
|
DOI |
https://doi.org/10.5281/zenodo.5575715 |
|
persistent identifier |
https://treatment.plazi.org/id/03CE87AD-FFED-5F49-FE4F-FABCFD145725 |
|
treatment provided by |
Jeremy |
|
scientific name |
Sesieutes Simon, 1897 |
| status |
|
Genus Sesieutes Simon, 1897 View in CoL View at ENA
Sesieutes Simon, 1897: 153 View in CoL . Deeleman-Reinhold, 2001: 446 Type species
Sesieutes lucens Simon, 1897 View in CoL , by monotypy.
Diagnosis
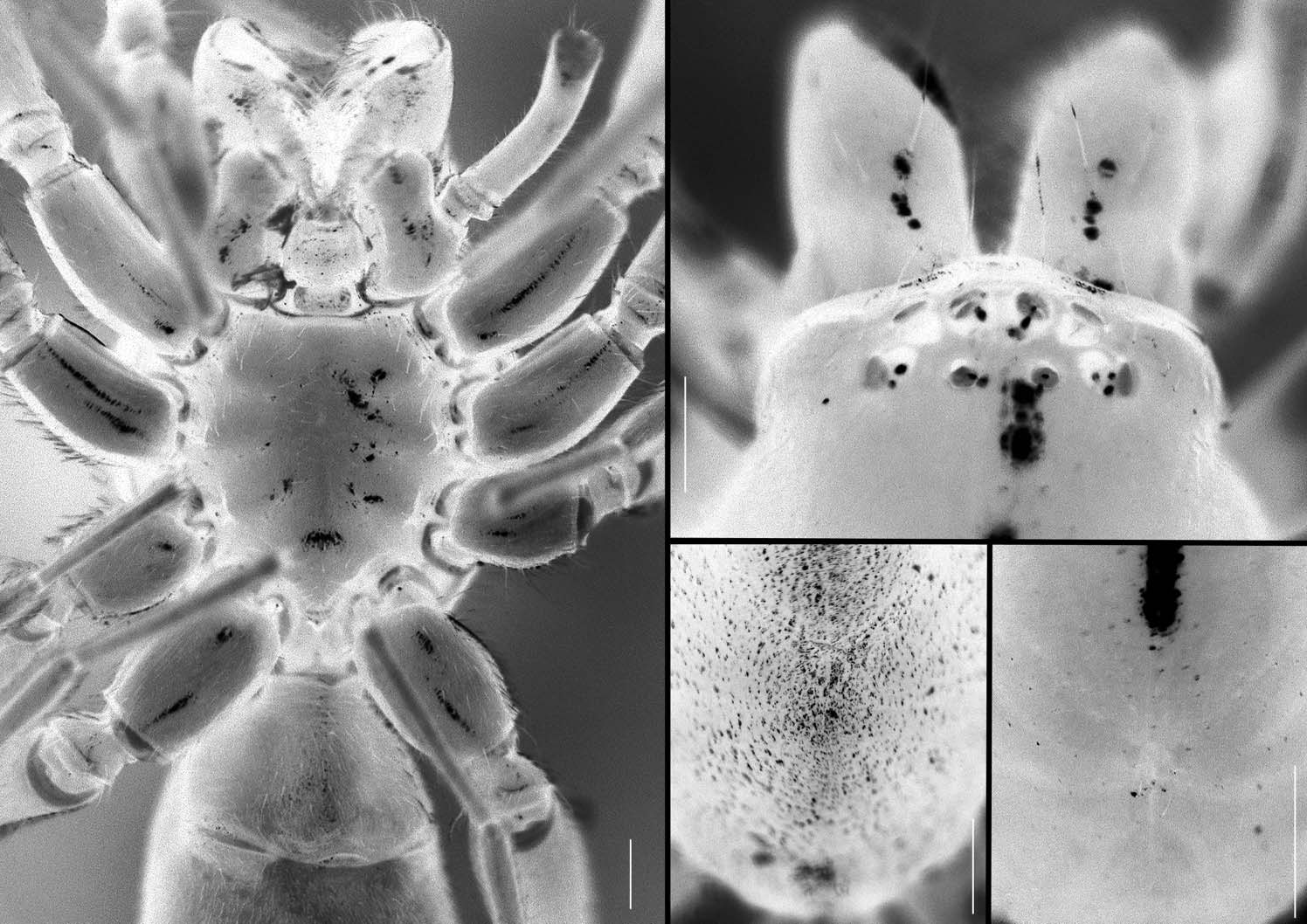
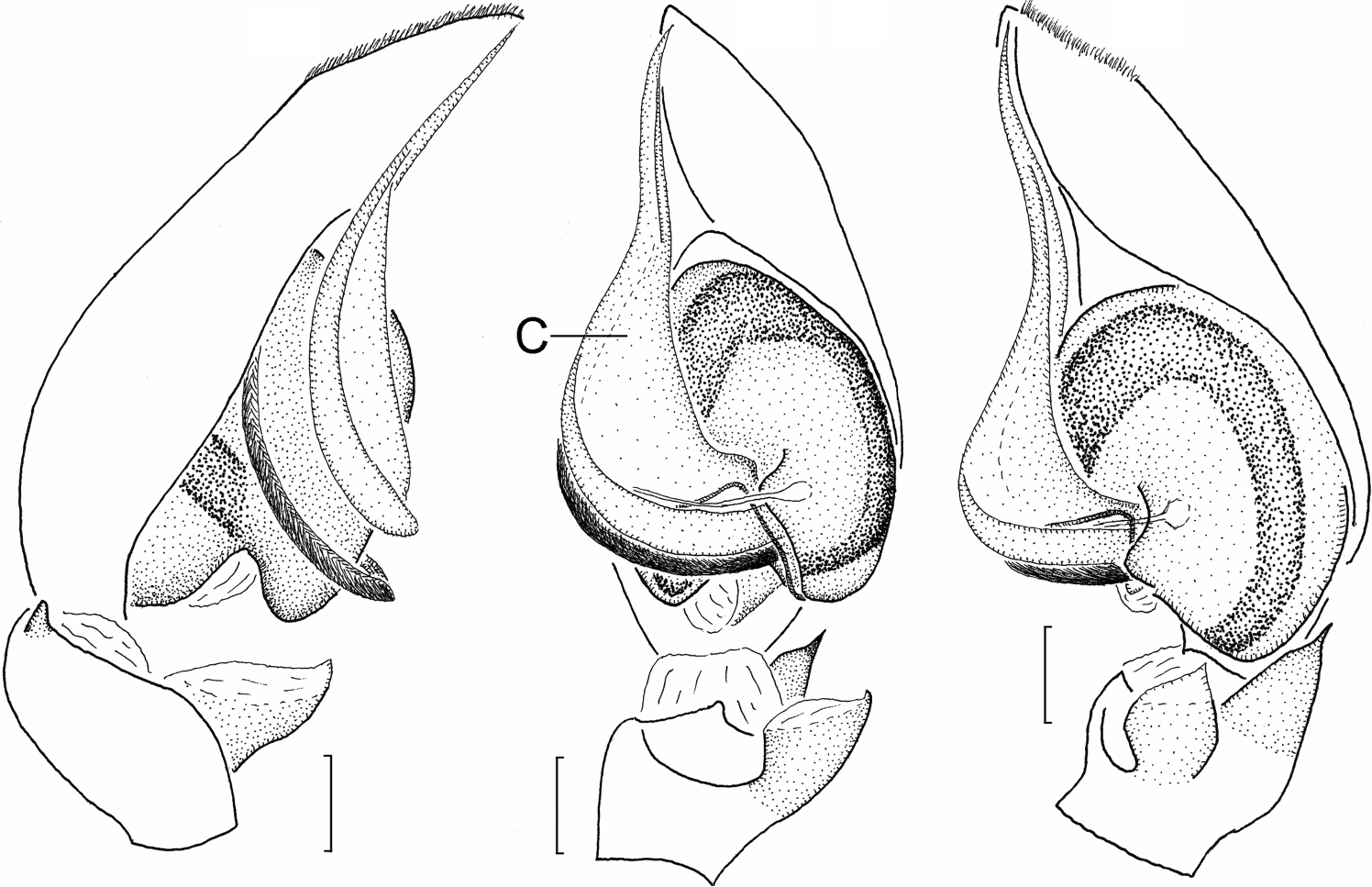
Representatives of Sesieutes can be distinguished from South East Asian phrurolithines by a combination of the following characters: the strongly attenuated carapace is relatively low, with an indistinct fovea ( Figures 2C, D View Figure 2 ), a clypeal lobe ( Figure 2B View Figure 2 ), and short, straight eye rows; the PME are smallest and widely separated, with distance between PME–PME subequal to PME–PLE ( Figure 2B View Figure 2 ); the opisthosoma is covered with sharply defined edge dorsal scutum in both sexes ( Figure 1 View Figure 1 ). Males can be further recognized by: male palpal tibia consists of a welldeveloped prolateral lobe ( Figure 4B, PE View Figure 4 ), a truncate, partially membranous proximal RTA ( Figure 4A, pRTA View Figure 4 ), and a heavily sclerotized distal RTA of variable shapes and sizes ( Figure 3 View Figure 3 ); the elongated, filiform embolus is curved behind apical part of tegulum, its apex always aligned with lanceolated conductor and spiniform tegular apophysis ( Figure 5A–C View Figure 5 ). Females can be recognized by the large copulatory orifices on the heavily sclerotized epigastric scutum ( Figure 5D View Figure 5 ) leading to the elongated, intertwined, mid-longitudinally arranged insemination ducts and the elliptical, terminal spermathecae ( Figure 5E View Figure 5 ).
Description
Prosoma ( Figure 1 View Figure 1 ) ovoid in dorsal view, strongly attenuated in front; widest between coxae II and III; relatively low, in profile highest mid distance between PME and fovea, gradually sloping posteriorly; carapace integument from smooth and shiny to strongly granulated, colour pale yellow to bright orange and dark brown. Fovea indistinct, represented by shallow depression ( Figure 2C,D View Figure 2 ). Ocular area and clypeus with few setae, other areas devoid of setae except those situated on elevated granules; clypeus strongly rebordered, with small clypeal lobe ( Figure 2B View Figure 2 ); from above, both anterior and posterior eye rows straight; both eye rows short, never occupying entire attenuated area ( Figure 2B View Figure 2 ); AME circular, light, largest; PME smallest, oval, pearly white; ALE and PLE circular, light; anterior eyes contiguous; PME separated by twice their diameter, about the same distance to PLE; ALE and PLE separated by more than their diameter; PER slightly wider than AER, both almost parallel; MOQ longer than wide, wider in front than behind; clypeal height subequal to AME diameter. Chelicerae convex anteriorly, with conspicuous condyles, frontal surface lined with few bristles, devoid of macrosetae; fang furrow with three to four promarginal teeth and one to two retromarginal denticles. Chilum trapezoidal, heavily sclerotized. Labium ( Figure 2A View Figure 2 ) longer than wide, apex straight, invaginated at posterior lateral margins. Gnathocoxae rectangular, with oblique depressions (more prominent in males), each with anteromedian scopula arising from white apical area ( Figure 2A View Figure 2 ). Sternum scutiform, longer than wide, slightly convex, with shallow radiating grooves and few bristles; anterior margin of sternum straight, with shallow median invagination; lateral margin of sternum bearing short, triangular, sclerotized extensions fitting to coxae; spaces between coxae with chitinized strips extending dorsally into pleural membrane, connecting between sternum and carapace; posterior margin of sternum strongly produced between coxae IV, with enlarged chitinized plate ( Figure 2A View Figure 2 ). Leg formula 4123; legs brownish orange; coxae I lengthened, coxae III shortest; trochanters not notched; femora I always with apico-prolateral spine; anterior tibiae not ventrally flattened, with five to seven pairs of ventral spines and single unpaired prolateral or mid-ventral spine; anterior metatarsi with four to six pairs of ventral spines, posterior metatarsi with bush of distoventral hairs; tarsi spineless, with two pectinate claws carrying three to eight denticles, lacking distinct claw tufts; trichobothria long, in two rows on tarsi, one row on metatarsi and tibiae, increasing in length distally.
Opisthosoma elongate-ovoid, dorsoventrally flattened, dark grey dorsally, greyishwhite ventrally; dorsal pattern consisting of pale, narrowed chevrons situated underneath dorsal scutum ( Figure 1O View Figure 1 ). Dorsal scutum with sharply defined edge, present in both sexes; elliptical, covering three-quarters of opisthosomal length or entirely in males; from short, semi-circular plate situated anteriorly to elliptical plate occupying three-quarters of opisthosomal length in females. Epigastric region entirely covered with scutum ( Figure 2A View Figure 2 ), extending anteriorly to form groove collar ( Figure 1N View Figure 1 ), heavily sclerotized, tightly fused with dorsal scutum. Ventral scutum absent. Postgenital scuta ( Figure 2A View Figure 2 ) rectangular or semi-circular, situated posterolaterally, surrounding lung slits but clearly separated from epigastric scutum. Spinnerets two-segmented; ALS conical, separated by slightly less than their diameter, distal segment short; PMS small, slender in males, flattened and elongated in females; PLS with short distal segment.
Male palpal femur with modified ventral surface in some species. Palpal tibia with well-developed prolateral excrescence ( Figures 3 H, 4 B, 5 B). Retrolateral tibial apophysis consisting of basal and distal parts; basal RTA represented by elevated ridge and partially membranous anterior surface ( Figure 5A–C View Figure 5 ) or apex ( Figure 4A–C View Figure 4 ); distal RTA, if present, heavily sclerotized ( Figure 4 View Figure 4 A). Cymbium apically with midlongitudinal patch of hairs ( Figure 3H–L View Figure 3 ). Tegulum pyriform, with distinct invagination accommodating base of lanceolated, semi-transparent conductor. Embolic base usually originating on apico-prolateral surface of tegulum ( Figure 5A,B View Figure 5 ); embolus filiform, elongated, curving behind tegulum ( Figure 5A–C View Figure 5 ); tegular apophysis hyaline, filiform, originating near base of conductor ( Figure 9C View Figure 9 ), aligned with embolic tip and conductor, their apices usually reaching apex of cymbium ( Figure 4A View Figure 4 ).
Epigynal region heavily sclerotized, with two large copulatory orifices and narrowed, introductory grooves ( Figure 5D View Figure 5 ); these presumably responsible for guiding elongated embolus and sclerotized conductor towards anteriorly located copulatory orifices during copulation; this region usually filled with dark secretory substance. Internal genitalia ( Figures 4 E, 5 E) consisting of transparent, medially situated proximal ducts (PD), strongly convoluted insemination ducts (ID), and laterally situated spermathecae (S). Spermathecae elliptical or reniform, not being separated structures but terminal enlargement of insemination ducts, usually with gland pores.
Included species
Sesieutes aberrans sp. nov., S. abruptus sp. nov., S. apiculatus sp. nov., S. bifidus sp. nov., S. borneensis Deeleman-Reinhold, 2001 , S. bulbosus Deeleman-Reinhold, 2001 , S. emancipatus Deeleman-Reinhold, 2001 , S. lucens Simon, 1897 , S. minor Deeleman-Reinhold, 2001 , S. minuatus sp. nov., S. nitens Deeleman-Reinhold, 2001 , S. scrobiculatus Deeleman-Reinhold, 2001 .
Possibly misplaced species
Four species are misplaced in Sesieutes and will be transferred to Jacaena Thorell, 1897 in a separate paper once that genus is redefined and revised. These taxa are S. erawan Deeleman-Reinhold, 2001 , S. schwendingeri Deeleman-Reinhold, 2001 , S. thakek Jäger, 2007 and S. zhui Zhang and Fu, 2011 .
Distribution
Indo-Burmese and Sundaland of the Oriental Region.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Sesieutes Simon, 1897
| Dankittipakul & Deeleman-Reinhold 2013 |
Sesieutes
| Simon 1897: 153 |
Sesieutes lucens
| Simon 1897 |