
Amphoriscus chrysalis ( Schmidt, 1864 )
|
publication ID |
https://doi.org/10.11646/zootaxa.5061.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:EFE2D12B-3F02-4831-A913-DEFE1F9A0C92 |
|
DOI |
https://doi.org/10.5281/zenodo.5642236 |
|
persistent identifier |
https://treatment.plazi.org/id/03CEAF23-A466-FF85-FF36-ED9CF017FCCD |
|
treatment provided by |
Plazi |
|
scientific name |
Amphoriscus chrysalis ( Schmidt, 1864 ) |
| status |
|
Amphoriscus chrysalis ( Schmidt, 1864) View in CoL
( type species by subsequent designation; Dendy & Row,1913)
Citations and synonymies: Ute chrysalis Schmidt 1864: 23 (original description); Amphoriscus chrysalis Haeckel 1870: 177 ; Sycilla chrysalis Haeckel 1872: 256 ; Sycurus chrysalis Haeckel 1872: 256 (variation of Sycilla chrysalis ); Amphoriscus chrysalis Poléjaeff 1883: 7 ; Dendy & Row 1913: 782; Breitfuss 1935: 29; Burton 1963: 535; Vacelet 1981: 165; Radolović et al. 2015: 305; Klautau, Cavalcanti & Borojevic 2017: 105; Cóndor-Luján et al. 2019: 1825.
Type material: Unknown.
Type locality: Lesina, Lissa, Adriatic Sea.
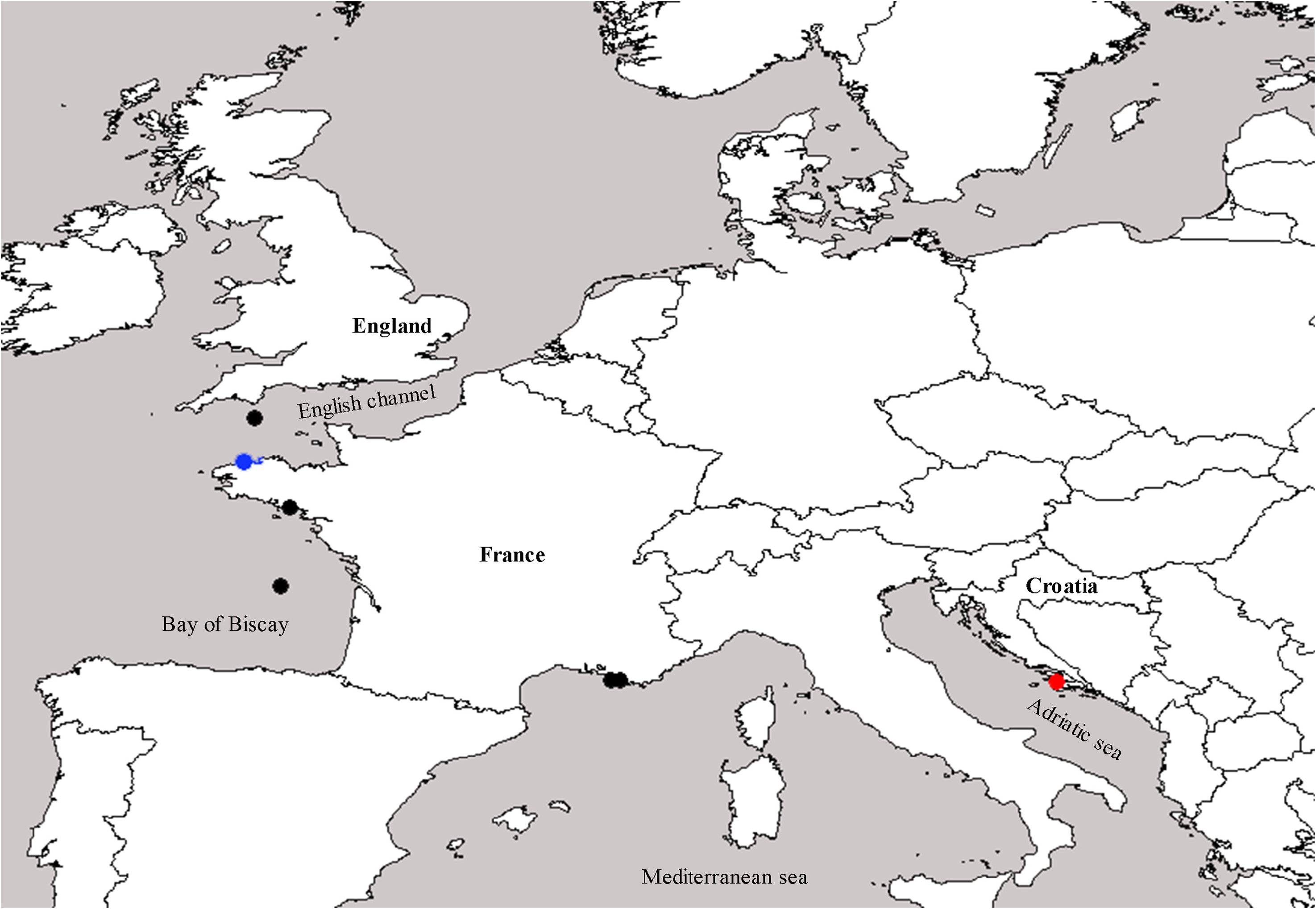
Analysed material: LBIM 1968-557 L ( Glenans, France; four slides containing sections of the skeleton) . LBIM 1968-556 L ( Marseille, France; three slides containing sections of the skeleton) . LBIM 1968-327 ( Roscoff, France; four slides containing sections of the skeleton) . LBIM 1968-355 ( Roscoff, France; two slides containing sections of the skeleton) . LBIM 1968-326 ( Roscoff, France; seven slides containing sections of the skeleton and dissociated spicules) . LBIM 1968-280 ( Roscoff, France; one slide containing dissociated spicules) . BMNH 1954.8 .12.195 ( Plymouth; M.B. L. collection; one slide containing sections of the skeleton) . BMNH 195512.13 .9 ( Plymouth; R.W.H. collection; one slide containing sections of the skeleton; not A. chrysalis — see below). SME-500 ( Biscay Bay , France; one specimen and one slide containing sections of the skeleton) .
Morphology: Colour in spirit varies from yellowish-white to yellowish-brown ( Burton 1963). The specimen SME-500 has a tubular shape and measures ca. 4 mm x 1 mm ( Fig. 1A View FIGURE 1 ). According to Schmidt (1864), the type had a peduncle, which is not visible in SME-500. Smooth surface and a single osculum without fringe of trichoxeas. Syconoid aquiferous system. Abundant larvae were observed in the choanosome of LBIM 1968-556 L ( Figs. 1B, D, E View FIGURE 1 ).
Anatomy: Skeleton typical of the family Amphoriscidae ( Fig. 1B View FIGURE 1 ). The cortical region is formed mainly by giant and abundant tetractines and a few tangential triactines, pierced by tufts of trichoxeas ( Fig. 1C View FIGURE 1 ). Abundant subatrial tetractines and triactines point their unpaired actine towards the cortex ( Fig. 1E View FIGURE 1 ). The atrial skeleton is formed exclusively by tetractines, and these have a very long apical actine ( Fig. 1F View FIGURE 1 ). Some lacunae occur along the subcortical and subatrial regions, mainly in the specimen LBIM 1968-556 L, possibly due to fixation and/or the occurrence of tissue retraction over time ( Fig. 1D View FIGURE 1 ).
Fusiform diactines were found in the spicules slides of LBIM 1968-326, but these were not observed in the sections of the skeleton of the same specimen and were not found in any other analysed material. In addition, BMNH 1955.12.13.9 differs from the remaining analysed samples in its skeletal composition: cortical tetractines, subatrial triactines, and atrial tetractines, and thus cannot be classified as A. chrysalis (see Remarks).
Spicules: Among the analysed material, slides containing dissociated spicules were available only for LBIM 1968-326. The values shown here therefore only reflect this sample.
Cortical tetractines ( Fig. 2A View FIGURE 2 ): Conical with blunt tips. The paired actines are curved, and one of the tips is broken in most of the spicules (118.8– 298.3 ±90.5–520.2/ 23.8– 38.9 ±7.9–53.1 μm). The unpaired actine is smaller or the same size as the paired actines (106.1– 203.6 ±107.4–485.1/ 22.2– 41.5 ±8.0–58.6 μm). The apical actine is straight and longer than the other actines (281.5– 421.6 ±87.2–636.2/ 23.8– 39.8 ±7.7–53.7 μm).
Cortical triactines ( Fig. 2B View FIGURE 2 ): Conical with blunt tips, less abundant than the cortical tetractines. Paired actines are slightly curved (246.2– 360.5 ±66.8–466.0/ 22.3– 35.6 ±6.3–45.2 μm). The unpaired actine is straight, sometimes slightly lanceolate (241.7– 468.8 ±140.7–673.9/ 23.1– 37.2 ±8.1–52.6 μm).
Subatrial triactines and tetractines ( Figs. 2C, D View FIGURE 2 ): Cylindrical to slightly conical and blunt. The paired actines are short and straight (triactines: 102.1– 145.0 ±23.3–185.3/ 6.2– 10.2 ±1.9–13.6 μm; tetractines: 92.9– 139.7 ±45.5–253.1/ 6.3– 8.5 ±2.0–11.4 μm). The unpaired actines are straight and long (triactines: 232.5– 340.4 ±49.6–435.3/ 8.5– 13.0 ±2.4–17.4 μm; tetractines: 142.6– 271.3 ±67.6–350.8/ 8.0– 9.6 ±1.2–11.1 μm). The apical actine of the tetractines is short and curved (20.3– 31.0 ±14.3–68.6/ 5.3– 7.6 ±1.7–10.1 μm).
Atrial tetractines( Fig.2E View FIGURE 2 ):Cylindrical with sharp tips.The paired actines are long and curved (100.3– 173.0 ±49.3– 315.4/ 6.0– 14.1 ±15.9–97.1 μm). The unpaired actine is straight (62.5– 169.3 ±95.9–445.6/ 4.9– 12.3 ±3.7–21.2 μm). The apical actine is straight and long. It is generally twice as large as the paired actines (102.2– 271.7 ±126.4–527.2/ 7.9– 11.9 ±2.6–17.8 μm).
Remarks: According to the original description ( Schmidt 1864), Amphoriscus chrysalis is comprised only of tetractines (the presence of only this spicule category and the external morphology are the only characters mentioned by the author). The material analysed here has cortical and subatrial triactines, differing from the original description. Haeckel (1872) also described A. chrysalis as having only tetractines, though subatrial (and possibly cortical) triactines similar to those we observed were illustrated in his work (Plate 43, figures 2 and 3). It is not clear if Haeckel analysed the same material described by Schmidt (1864), but the short original description lacking either spicule measurements or illustrations of the spicule types and skeleton show the importance of Haeckel’s (1872) work for the recognition of A. chrysalis . The difficulty in recognising this species is also caused by a lack of type material. Our results and the figure provided by Haeckel (1872) thus allow us to state that A. chrysalis has triactines. We were also able to recognise the main feature of A. chrysalis reported by Haeckel (1872) in the samples analysed here: the long apical actine of the atrial tetractines (>100 μm). This feature was important for supporting our decision to identify some samples as A. chrysalis sensu Haeckel (1872) .
Among the analysed samples, BMNH 1954.8.12.195 and BMNH 1955.12.13.9 had been listed by Burton (1963: 634) under the name A. chrysalis . Our results indicate that the latter does not belong to A. chrysalis due to its different skeletal composition. Here, we allocate it in A. decennis sp. nov., while BMNH 1954.8.12.195 remains A. chrysalis . Some of the samples analysed in this study had previously been identified as A. chrysalis by Borojevic et al. (1968), namely: LBIM 1968-557, LBIM 1968-556 L, LBIM 1968-327, LBIM 1968-355, LBIM 1968-326, and LBIM 1968-280. The list of species from Roscoff was provided without morphological details, meaning that these specimens are described for the first time in this study.
Compared to the other species of the genus, the most similar to A. chrysalis is A. elongatus . However, the main difference between them is the abundance of tetractines: the former has abundant tetractines in the subatrial region, while in A. elongatus tetractines were observed to be rare. Additionally, the actines of the subatrial triactines of A. chrysalis are more conical and bear blunt tips, unlike A. elongatus , which has more cylindrical actines and with sharp tips.
Distribution: Amphoriscus chrysalis is one of the few species of the genus that has several records besides its type locality. It has been described in the Adriatic and Celtic Seas ( Schmidt 1864; Borojevic et al. 1968) and is reported here in the Mediterranean part of France and the Bay of Biscay ( Fig. 3 View FIGURE 3 ). The corresponding marine ecoregions of the world (MEOW) are the Celtic Seas, South European Atlantic Shelf, Adriatic Sea, and Western Mediterranean ( Spalding et al. 2007).


| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Calcaronea |
|
Order |
|
|
Family |
|
|
Genus |
Amphoriscus chrysalis ( Schmidt, 1864 )
| Chagas, Cléslei & Cavalcanti, Fernanda F. 2021 |
Ute chrysalis
| Schmidt 1864 |