
Anguinaria truncata Landsborough, 1852: 288
|
publication ID |
https://doi.org/10.1206/0003-0090(2002)270<0001:NABFTV>2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/03D1878C-1947-FFB8-FD50-C17BFC12C729 |
|
treatment provided by |
Felipe |
|
scientific name |
Anguinaria truncata Landsborough, 1852: 288 |
| status |
|
Anguinaria truncata Landsborough, 1852: 288 . Aetea truncata: Gautier, 1962: 29 View in CoL . Hayward and
Ryland 1998: 104.
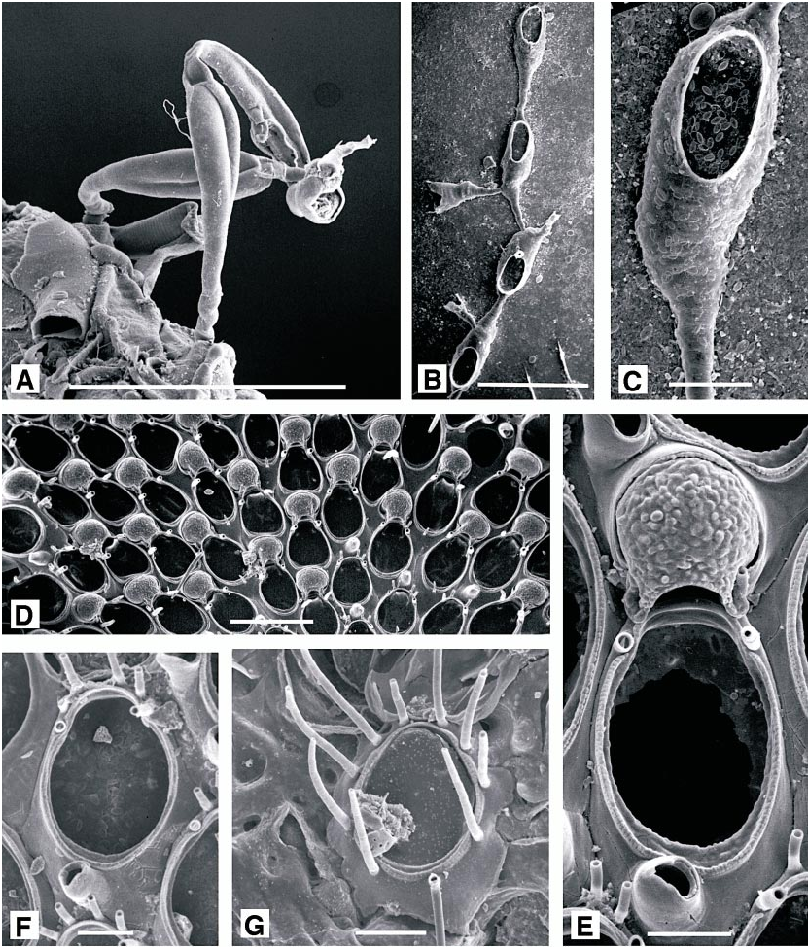
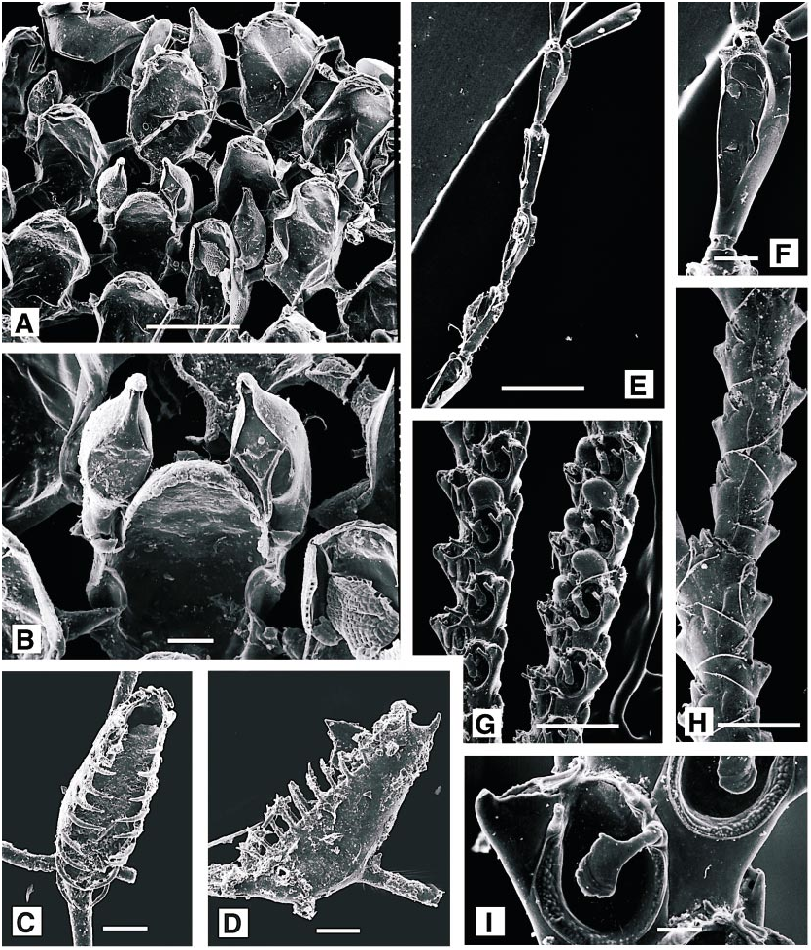
DESCRIPTION (AMNH 892, 893; CMRR 2204): Colonies white, uniserial, continuously or intermittently encrusting. Adnate stoloniform portion of autozooid particularly slender, as is basal dilatation of erect distal portion. Relatively short distal portion of autozooid finely and densely punctate, lacking any annulations. Colonies collected 26 May 1988 offshore from Rt Kriz had irregular unattached growth form reported in other species of Aetea ( Hayward and Ryland, 1998) when associated with hydroid and bryozoan turfs. They were loosely intergrown with a tuft of Savignyella lafontii . Stoloniform portions were up to 0.125 mm long but just 0.025 mm wide, broadening abruptly to 0.07 mm at base of distal portion, which was reflected at an angle of about 120° to long axis of stoloniform portion. Distal portion, accommodating the polypide, was 0.45–0.5 mm long, cylindrical proximally and slightly broadened to a maximum width of 0.12 mm at midlength. Opesia occupying about half of total length of this part of autozooid. Distally, autozooid abruptly truncate, bearing a terminal operculum. Branching varied: new stoloniform buds arising most frequently from basal dilatation of autozooid, either basally or laterally, or from basal (i.e., abfrontal) surface of distal portion of autozooid, or even from various points on stolonifom parts of autozooids.
Tentacles clear, 11–12, usually 12; lophophore broadly conical, radially symmetrical.
REMARKS: Geometry of adnate and erect portions of autozooids, together with punc tuation and lack of annulations suffice to distinguish this species from all other Mediterranean species of Aetea . Balduzzi et al. (1991) noted that the superficially similar A. lepadiformis Waters has a very few, but distinct annulations immediately proximal to the opesia
OCCURRENCE: This species was common to abundant at most stations where it occurred, encrusting dead shell and as an epizooite, especially of other bryozoans.
DISTRIBUTION: Aetea truncata has been reported from temperate and tropical seas throughout the world but, especially following the work of Balduzzi et al. (1991), it must be considered probable that such a wide Distribution encompasses more than one species.
MEASUREMENTS (POLYPIDE): IH 46 ± 36 µm, 20–120 (4, 17), LD 421 ± 57, 280–530 (4, 29), MD 21.7 ± 2.5, 20–25 (3, 9), TL 299 ± 55, 160–440 (4, 27).
SUBORDER SCRUPARIINA SILÉN, 1941 View in CoL SUPERFAMILY SCRUPARIOIDEA BUSK, 1852 View in CoL FAMILY SCRUPARIIDAE GRAY, 1848 View in CoL GENUS SCRUPARIA OKEN, 1815 View in CoL
Scruparia ambigua ( d’Orbigny, 1841) View in CoL Figure 6A View Fig
Eucratea ambigua d’Orbigny, 1841 : pl. 3, figs 13–17.
Scruparia ambigua: Hayward and Ryland, 1998: 108 View in CoL .
DESCRIPTION (AMNH 894): Colony erect, branching, uniserial chains of hornshaped autozooids, budded from encrusting chains of identical autozooids. Frontal surface of autozooid with an oval opesia occupying half its length, bounded by a thin raised rim. Embryos brooded in dimorphic zooids with a distinctive bivalved brood chamber (absent from present material).
OCCURRENCE: A portion of one colony, comprising two encrusting autozooids and two erect shoots, of three and one complete autozooids, encrusting Cellaria fistulosa ( Linnaeus, 1758) .
DISTRIBUTION: This rather inconspicuous species is widely distributed throughout the Mediterranean, and has been reported from temperate and tropical seas worldwide. It is often epifaunal in habit, but is rather inconspicuous and frequently overlooked.
SUBORDER MALACOSTEGA LEVINSEN, 1902 SUPERFAMILY MEMBRANIPOIDEA BUSK, 1852 FAMILY ELECTRIDAE D’ORBIGNY, 1851 View in CoL GENUS PYRIPORA View in CoL D’ORBIGNY, 1852
Figure 6B, C View Fig
DESCRIPTION (AMNH 895): Colony encrusting, runnerlike, inconspicuous. Autozooids in uniserial chains, branching irregularly; clavate, with a slender, filiform proximal portion broadening at about half total length to an oval distal portion. Calcification gymnocystal, smooth; opesia oval, occupying threequarters length of distal portion, bordered by a very narrow, indistinct cryptocystal rim. No spines or polymorphs. Each autozooid giving rise to a single distal bud, and may also produce paired lateral buds, allowing branching of the chains.
REMARKS: The single colony of Pyripora in our material differs from P. catenularia ( Fleming, 1828) , the species previously reported from the northeastern Atlantic and Mediterranean, in lacking a welldeveloped cryptocystal rim around the opesia, having a proportionally longer cauda, and having proximolateral rather than more distolateral budding.
MEASUREMENTS (SKELETAL): OpL 155 ± 12 µm, 29–168 (1, 10), OpW 81 ± 8, 70–93 (1, 10), ZL 548 ± 50, 480–621 (1, 10), ZW 155 ± 9, 141–170 (1, 10).
SUBORDER FLUSTRINA SMITT, 1867b
SUPERFAMILY CALLOPOROIDEA NORMAN, 1903 View in CoL
FAMILY CALLOPORIDAE NORMAN, 1903 View in CoL
GENUS CALLOPORA GRAY, 1848 View in CoL
Callopora dumerilii ( Audouin, 1826) View in CoL
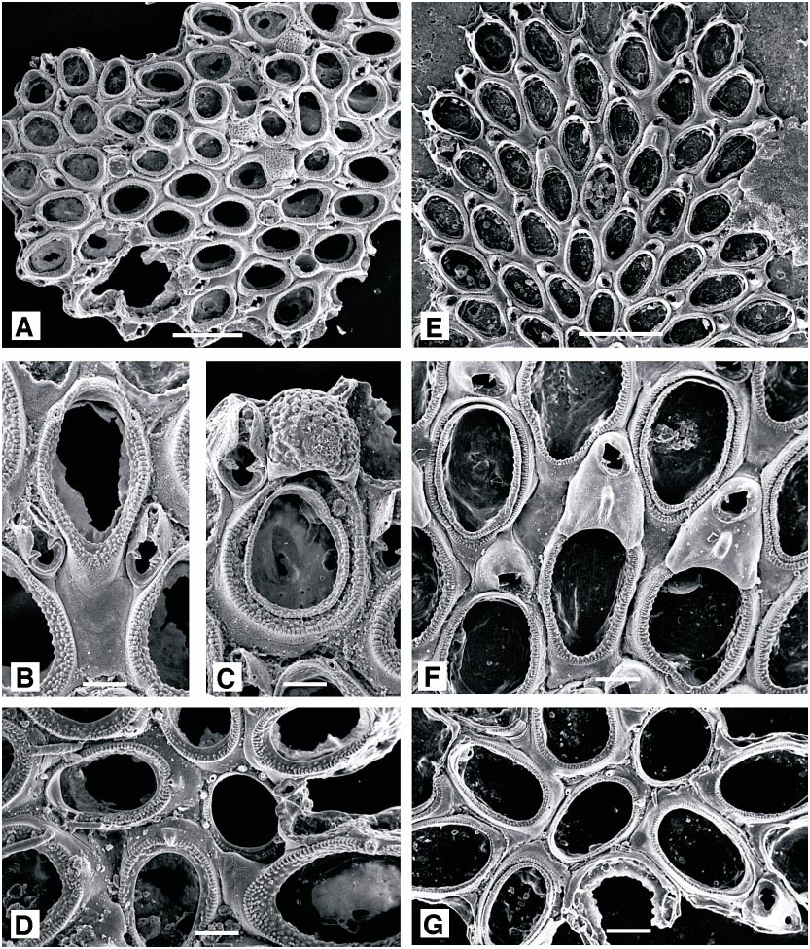
Figure 6D–G View Fig
Flustra dumerilii Audouin, 1826: 240 . Callopora dumerilii View in CoL ; Hayward and Ryland, 1998:
160.
DESCRIPTION (AMNH 896–898; CMRR 2205): Colonies encrusting, multiserial, unilaminar patches. Ancestrula approximately 225–250 µm long, with 10 peripheral spines;
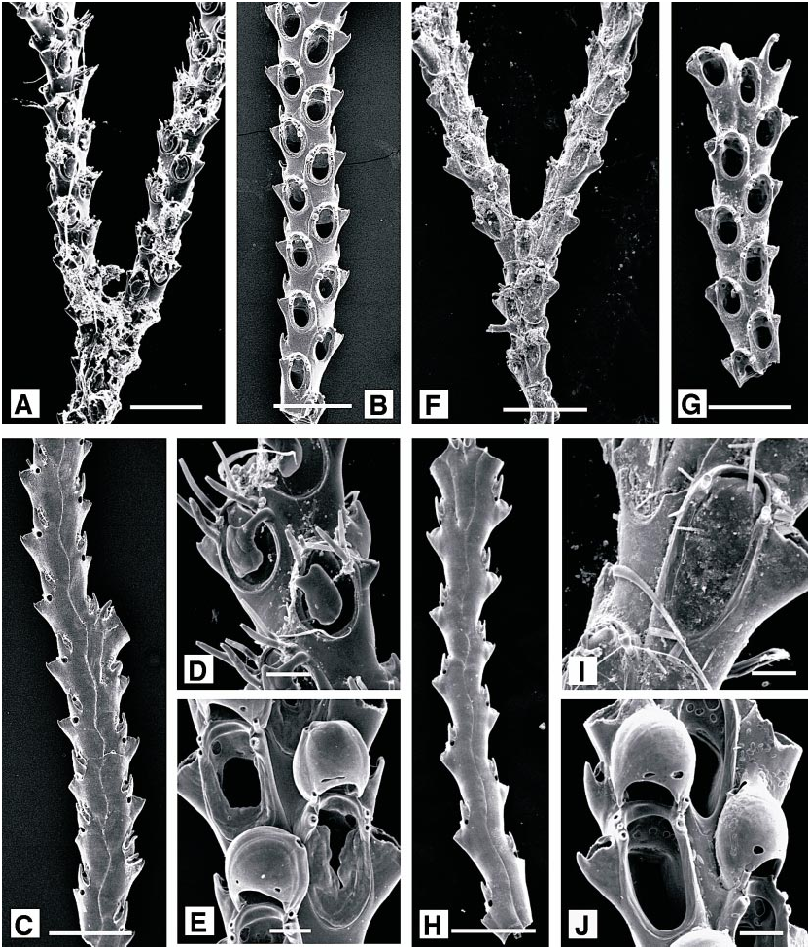
number of spines reduced and autozooids enlarged over about 5–8 generations. Autozooids elongate oval, broadest proximally, separated by distinct grooves. Opesia oval, bordered by a thin crenulate mural rim, enclosing a narrow shelf of finely granular cryptocyst, widest at the proximal end; gymnocyst smooth, imperforate. Four distal oral spines present, the proximalmost pair robust and conspicuous. A single adventitious avicularium (rarely two) present on the gymnocyst of most autozooids, with short, columnar cystid; rostrum acute to frontal plane, triangular, distally hooked, usually directed proximally; with a narrow cryptocyst bordering the proximal opesia, but no crossbar. Ovicell hyperstomial, prominent, more or less spherical, but narrowed towards arched aperture; ectooecium membranous except for a basal peripheral rim, entooecium coarsely nodular. Distal to an ovicell, avicularium displaced laterally, with a distolateral orientation.
Tentacles clear, 11–12; lophophores bellshaped, radially symmetrical.
DISTRIBUTION: Common throughout the temperate northeast Atlantic and Mediterranean, on stone, shell and other organic substrata, in shallow coastal habitats.
MEASUREMENTS (SKELETAL): DO 368 ± 52 µm, 285–497 (3, 30), OpL 294 ± 30, 222– 350 (3, 25), OpW 208 ± 16, 180–240 (3, 25), OvL 177 ± 17, 156–226 (3, 20), OvW 172 ± 21, 129–216 (3, 20), ZL 507 ± 37, 424–590 (3, 25), ZW 307 ± 25, 260–357 (3, 25). (POLYPIDE): IH 30–80 µm (1, 2), LD 373 ± 31, 340–400 (1, 3), MD 20 (1, 1), TL 287 ± 23, 260–300 (1, 3).
GENUS COPIDOZOUM HARMER, 1926 View in CoL
Copidozoum tenuirostre ( Hincks, 1880) View in CoL Figure 7A–D View Fig
Membranipora tenuirostre Hincks, 1880: 70 .
Copidozoum tenuirostre: Prenant and Bobin, 1966: 257 View in CoL . Hayward and Ryland, 1998: 180.
DESCRIPTION (AMNH 899, 900): Colonies encrusting, multiserial, unilaminar. Autozooids elongate oval, distinctly separated by wellmarked grooves. Frontal surface of autozooid almost entirely membranous; gymnocyst reduced to a smooth, proximal and proximolateral border; cryptocyst developed as a coarsely granular rim, narrowed slightly at distal end of autozooid, opesia constituting almost threequarters total autozooid length. Spines absent, except in first three or four astogenetic generations. Avicularia interzooidal, about half length of autozooid; mandible slender, distally directed, lodged within a narrow, parallelsided rostrum formed from incurved gymnocystal calcification, pivoting on two pronounced condyles, proximal to which there is a broad shelf of cryptocyst. Ovicell longer than wide, domed, with a distinctive coarsely nodular surface. Ancestrula oval, with minimal development of cryptocyst, bordered by (?) eight slender spines; autozooids of the first three or four astogenetic generations occasionally with single spines proximally, and two or more distally, all developed on the gymnocyst.
DISTRIBUTION: On stones and shells, in shallow coastal habitats; C. tenuirostre is widely distributed in the Mediterranean and north to the western English Channel.
MEASUREMENTS (SKELETAL): AL 229 ± 31 µm, 167–298 (2, 15), AW 123 ± 14, 94–155 (2. 15), DO 376 ± 63, 256–490 (3, 30), OpL 315 ± 29, 268–381 (3, 30), OpW 205 ± 24, 162–256 (3, 30), OvL 177 ± 9, 162–190 (2, 7), OvW 190 ± 14, 176–217 (2, 7), ZL 484 ± 56, 391–560 (3, 30), ZW: 322 ± 26, 279– 379 (3, 30).
GENUS ELLISINA NORMAN, 1903
Ellisina gautieri Fernández Pulpeiro and
Reverter Gil, 1993
Figure 7E–G View Fig
Ellisina gautieri Fernández Pulpeiro and Reverter Gil, 1993: 98 . Hayward and Ryland, 1998: 192. DESCRIPTION (AMNH 901): Colonies encrusting, multiserial, unilaminar patch. Autozooids oval, separated by distinct grooves. Frontal surface almost entirely membranous, the extensive opesia bordered by a very narrow rim of granular cryptocyst, and with a minimal area of smooth gymnocyst just visible at proximal end of autozooid. No spines. Avicularia interzooidal, budded from distal pore chamber of most autozooids; cystid rounded quadrangular, ca. 0.125 mm long, rostrum triangular, acute to frontal plane, directed distolaterally. Two or more avicularia may occur at the division of autozooid rows, or where regular spacing of autozooids is disturbed. Ovicell slightly longer than wide, domed, recumbent on substratum between distally succeeding autozooids; smooth and imperforate, with a distinct, longitudinal frontal ridge and intimately associated with distal avicularium; closed by zooidal operculum.
REMARKS: This species was first noted from Marcella by Gautier (1962), as E. cf. levata , and its identity was only recently established by Fernández Pulpeiro and Revert er Gil (1993).
DISTRIBUTION: E. gautieri is known from the Atlantic coast of northwest Spain and from the western English Channel; the single small colony described here represents its first record from the eastern Mediterranean.
MEASUREMENTS (SKELETAL): AL 158 ± 22 µm, 131–198 (2, 20), AW 140 ± 20, 113– 184 (2, 20), DO 359 ± 36, 296–424 (2, 20), OpL 346 ± 27, 304–387 (2, 20), OpW 205 ± 15, 182–246 (2, 20), ZL 521 ± 42, 465– 596 (2, 20), ZW 296 ± 14, 274–325 (2, 20).
SUPERFAMILY FLUSTROIDEA FLEMING, 1828 View in CoL FAMILY FLUSTRIDAE FLEMING, 1828 View in CoL GENUS CHARTELLA GRAY, 1848 View in CoL
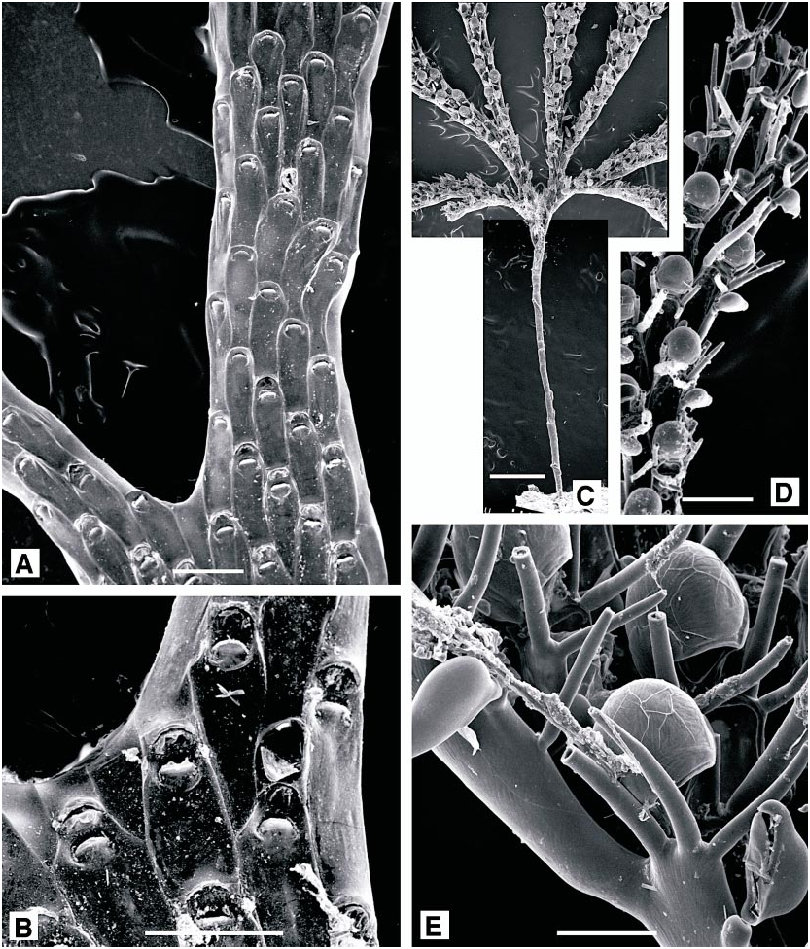
Chartella tenella ( Hincks, 1887) View in CoL Figure 8A, B View Fig
Flustra tenella Hincks, 1887: 313 .
Chartella tenella: Gautier, 1962: 48 View in CoL .
Terminoflustra tenella: Prenant and Bobin, 1966: 191 .
DESCRIPTION (AMNH 902; CMRR 2206): Colonies erect, flabellate, bilaminar, lightly calcified, to 20 mm in present material; dividing irregularly at varying intervals, developing a narrowly lobed, frondose form. Autozooids rectangular, flat, 0.65 X 0.25 mm. Frontal surface entirely membranous, vertical walls lightly calcified; a short, thin and very inconspicuous spine present at each distal corner. Avicularia vicarious, at bifurcation of autozooid rows; cystid rectangular, 0.25 X 0.2 mm, thickly calcified, with semielliptical rostrum slightly acute to frontal plane, directed distally or just oblique to distal axis. Kenozooids filling spaces between autozooids where bifurcating branches diverge and developed as tubular extensions at distal tips of some branches. Ovicell hemispherical, completely immersed, conspicuous through its white calcification.
Tentacles colorless, 13–15; lophophores bellshaped, radially symmetrical.
Light orange embryos present in material from locality 21, 15 June 1988.
DISTRIBUTION: Chartella tenella is an endemic Mediterranean species, common in shallow coastal habitats, and offshore to 200 m ( Prenant and Bobin, 1966). It has been widely reported throughout the western Mediterranean and the Adriatic.
MEASUREMENTS (SKELETAL): AL 314 µm (1, 1), AW 221 (1, 1), DO 444 ± 84, 301– 632 (2, 20), OL 76 ± 8, 62–92 (1, 10), OW 134 ± 8, 121–148 (1, 10), ZL 685 ± 81, 585–841 (2, 20), ZW 250 ± 36, 184–321 (2, 20). (POLYPIDE): IH 201 ± 74 µm, 60–340 (2, 24), LD 595 ± 94, 440–700 (2, 27), MD 20 (1, 1), TL 534 ± 62, 400–650 (2, 24).
SUPERFAMILY BUGULOIDEA GRAY, 1848 View in CoL FAMILY BUGULIDAE GRAY, 1848 View in CoL GENUS BUGULA OKEN, 1815 View in CoL
Bugula fulva Ryland, 1960 View in CoL Figure 8C–E View Fig
Bugula fulva Ryland, 1960: 86 View in CoL . Hayward and Ryland, 1998: 218.
DESCRIPTION (AMNH 903, 904; CMRR 2207): Colonies erect, branching, developing as stiff, open fans, to 10 mm in present material but recorded up to 30 mm. Branches unilaminar, dividing by regular dichotomy, consisting of two longitudinal autozooid series, increasing to four just prior to each dichotomy. Autozooids elongate, narrow, frontal dimensions ca. 0.6 X 0.2 mm, slightly tapered proximally; frontal surface entirely membranous. Usually three large spines at each distal corner, sometimes five on enclosed autozooids proximal to a dichotomy, the distalmost spine on the outer corner especially robust. Most autozooids with a lateral avicularium, attached on outer edge of frontal membrane just below proximalmost spine; length slightly greater than the autozooid width, but variable and locally less than width on some autozooids; beak gently downcurved. Ovicell large, globular, with a wide, arched aperture; entooecium with fine radiating striations. Ancestrula distinctive: very elongate, slender, trumpetshaped; tubular proximal portion to 2.5 mm in present material, with oval distal opesia bordered by six distal and three proximal spines, sometimes with an additional lateral pair between. Tubular rhizoids budded from proximal region of ancestrula anchor the colony.
REMARKS: Ancestrulae were present in every colony in 1997 and 1998 samples from the 35 m deep Cellaria meadow west of Banjole Island. They were remarkable for their length, resembling the basal kenozooids of colonies of Caulibugula , which caused this local population to be incorrectly identified in an ecological study ( McKinney and Jaklin, 2000). Both Prenant and Bobin (1966) and Hayward and Ryland (1998) figured the ancestrula of B. fulva , and the former noted that it might be very much longer than an autozooid, although none was reported to be as long as those described here. In other respects, however, the morphology of the ancestrulae from Rovinj is identical to that described previously and the exceptionally elongate, tubular proximal portion apparently reflects a phenotypic response to microhabitat characteristics. The pattern of autozooids at a bifurcation mostly conforms to the Type 5 of Harmer (1923), with zooid F enclosed proximal to the bifurcation, and zooid G adjacent to the axil. However, Hayward and Ryland (1998) note that this is inconstant and that some dichotomies within a colony may be achieved by simple splitting of the four autozooid series. In one of the Rovinj specimens this had occurred immediately distal to the ancestrula, and the first bud developed as another very elongate autozooid, reinforcing the specimen’s superficial resemblance to Caulibugula .
Tentacles clear, 14–15; lophophores bellshaped, all obliquely truncate.
DISTRIBUTION: Bugula fulva has been patchily recorded from the southwest British Isles and parts of the Mediterranean, low in the littoral zone and in shallow coastal waters. It is also associated with ports and harbors, and its wide occurrence in the western Atlantic, from Brazil to Maine, perhaps suggests an exotic origin as a fouling species.
MEASUREMENTS (CYSTID): AL 196 ± 19 µm, 170–235 (2, 19), DO 228 ± 34, 276– 405 (2, 20), OvL 212 ± 16. 185–242 (2, 20), OvW 194 ± 9, 180–209 (2, 20), ZL 577 ± 40, 500–660 (2, 20), ZW 178 ± 20, 130– 220 (4, 40). (POLYPIDE): IH 40 µm (1.10), LD 450 ± 54, 360–540 (1, 10), MD 20 (1, 1), TL 460 ± 51, 400–560 (1, 10).
FAMILY BEANIIDAE CANU AND BASSLER, 1927 View in CoL GENUS BEANIA JOHNSTON, 1840 View in CoL
Beania cylindrica ( Hincks, 1886) View in CoL
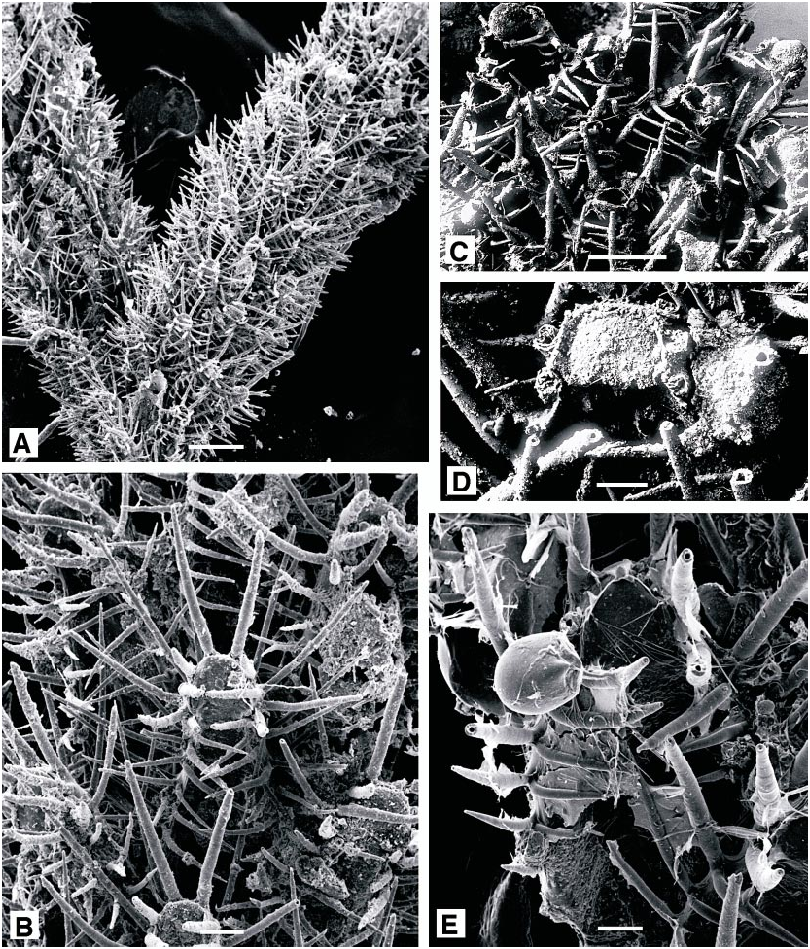
Figure 9A, B View Fig
Diachoris hirtissima forma cylindrica Hincks, 1886: 263 View in CoL .
Beania hirtissima: Gautier, 1962: 95 View in CoL . Prenant and Bobin, 1966: 557.
Beania hirtissima View in CoL forma cylindrica: Gautier, 1962: 96 View in CoL .
DESCRIPTION (AMNH 905; CMRR 2208): Autozooids elongate, narrow, commonly 1.0 X 0.25 mm; tapered proximally, truncate distally, with the terminal operculum orientated perpendicular to frontal plane. Frontal surface entirely membranous, bordered by long, thin spines: six erect spines evenly spaced around distolateral rim of operculum, eight to ten pairs of spines along margins of frontal membrane, slightly incurved; additional, equally long or longer, spines developed on lateral and basal surfaces of autozooids, aligned with those on frontal margins. Each autozooid linked to six neighbors by short, tubular connections: one proximal and two proximolateral, all closely spaced, and three on basal wall midway along the length of the autozooid.
Colonies are erect tufts, to 12 mm, loosely and irregularly branched. Primary branches of five to eight alternating autozooid series defining a convex frontal surface encompassing twothirds circumference of branch. Primary branches divide dichotomously at irregular intervals; minor branches of two to three autozooid series originating at axil of dichotomies or along branch margins. Abfrontal surface concave, with the space it encloses crossed by basal spines of autozooids. Septula on basal autozooid walls give rise to tubular kenozooidal rhizoids which pass proximally to attach colony to substratum.
Tentacles clear, 26; lophophores bellshaped, radially symmetrical.
REMARKS: Material described here conforms to the colony morphotype often denoted Beania hirtissima form or var. cylindrica . However, as discussed below, B. hirtissima has very different autozooid shape and orientation plus scattered vicarious avicularia, which are absent in B. cylindrica .
DISTRIBUTION: Beania cylindrica has been accorded a wide geographical Distribution, in shallow, warm temperate to tropical seas throughout the world, which possibly needs to be reexamined.
MEASUREMENTS (SKELETAL): DO 549 ± 92 µm,, 377–792 (2, 10), OL 139 ± 19, 117– 176 (2, 7), OW 138 ± 8, 120–149 (2, 6), ZL 993 ± 97, 900–1204 (2, 10), ZW 381 ± 34, 327–443 (2, 10). (POLYPIDE): IH 602 ± 80 µm, 500–700 (1, 10), LD 995 ± 72, 880– 1100 (1, 8), MD 40 (1, 1), TL962 ± 29, 920–1020 (1, 10).
Beania hirtissima ( Heller, 1867) View in CoL Figure 9C–E View Fig
Diachoris hirtissima Heller, 1867: 94 .
Diachoris hirtissima forma robusta Hincks, 1881: 133 .
Beania robusta: Gautier, 1962: 99 View in CoL . Zabala and Maluquer, 1988: 101.
LECTOTYPE (chosen here): UIIZ 34.
DESCRIPTION (AMNH 906; CMRR 2209): Colonies multiserial, unilaminar sheets of closely spaced, suberect autozooids, each linked to six neighbors by a ring of six short connecting tubes around proximal end. Autozooids broadest proximally, narrowest at the distal end; 0.8–1.0 mm long. Membranous frontal surface bordered by stout spines: three in a line along distal edge of the terminal operculum and two on each side, all especially long and straight, to 0.5 mm; five erect spines on each lateral margin, straight and successively shorter proximally, each with a shorter, thinner spine at its base, curving medially over frontal margin. Basal surface of colony with three to eight irregularly tapered and curved, thin processes on each autozooid.
Avicularia infrequent; pedunculate, attached to lateral wall of autozooid level with proximal corner of operculum; globose, with a broadly triangular, distally hooked rostrum.
REMARKS: We designate as lectotype Hell er’s specimen illustrated here as fig. 9C and 9D (UIIZ specimen 34, one of two specimens in the collection indicated as Heller’s types of the species). The lectotype has the features attributed to B. robusta , described originally from Algiers ( Hincks, 1881) as Diachoris hirtissima forma robusta .
DISTRIBUTION: Beania hirtissima has been accorded a wide geographical Distribution, in shallow, warm temperate to tropical seas throughout the world, which needs to be reexamined in light of the characteristics of the lectotype. As B. robusta , this rather substantial Beania has been reported from a number of localities in the northwestern Mediterranean, and also from Tunisia ( Gautier, 1962). It does not seem to have been recorded from east of the Adriatic.
MEASUREMENTS (SKELETAL): AL 216 ± 14 µm, 199–230 (1, 4), AW 170 ± 4, 166–176 (1, 4), DO 499 ± 65, 394–600 (1, 11), OL 161 ± 17, 141–194 (1, 10), OW 200 ± 13, 183–222(1, 10).
Beania magellanica ( Busk, 1852) View in CoL Figure 10A, B View Fig
Diachoris magellanica Busk, 1852: 54 .
Beania magellanica: Gautier, 1962: 97 View in CoL . Prenant and Bobin, 1966: 555.
DESCRIPTION (AMNH 907; CMRR 2210): Colonies forming repent sheets, supported above and anchored to substratum by short, tubular, kenozooidal rhizoids. Autozooids large, elongate oval, ca. 0.8 mm long in present material, membranous frontal surface with a smooth rim, lacking spines. Distal end of autozooid rounded, edge of the operculum fitting along distal rim; proximal end gently tapered. Autozooids disjunct, linked by tubular connecting processes, 0.2 mm long; each autozooid with six tubes: one midproximal, two proximolateral, and three distobasal, the latter equally spaced in an arc in distal third of the autozooid. Conspicuous, paired avicularia present, attached to lateral walls close to distal series of connecting tubes; pedunculate, with bird’s head form, rostrum tapered, curved, and distally hooked. At rest mandible lying in frontal plane of autozooid, with rostrum directed distally. Ovicell an inconspicuous, partly immersed, distal cap, not seen in present material.
Twentyseven tentacles (based on single count).
REMARKS: This is the most distinctive Beania species occurring in the Mediterranean region.
DISTRIBUTION: Widely distributed throughout the Mediterranean, in shallow coastal waters, most frequently on biogenic substrata. Beania magellanica has been reported from throughout the warm temperate to tropical Atlantic, as well as from the western Pacific.
MEASUREMENTS (SKELETAL): AL 439 ± 53 µm, 342–528 (3, 16), AW 151 ± 14, 118– 173 (2, 15), DO 586 ± 88, 438–689 (2, 15), OL 110 ± 12, 95–126 (1, 5), OW 158 ± 16, 143–184 (1, 5), ZL 713 ± 55, 634–799 (3, 13), ZW 440 ± 33, 373–478 (3, 13). (POLYP IDE): LD 1140 µm (1, 1), TL 980 (1, 1).
Beania mirabilis Johnston, 1840 View in CoL Figure 10C, D View Fig
Beania mirabilis Johnston, 1840: 272 View in CoL . Alvarez, 1990: 26. Hayward and Ryland, 1998: 244.
DESCRIPTION (AMNH 908; CMRR 2211): Colonies formed from uniserial chains of autozooids only lightly attached to substratum, often largely free. Each autozooid consisting of a filiform proximal portion comprising more than half its length, and a boatshaped distal portion, broadest proximally and tapered towards the terminal operculum. Calcification thin and translucent, the frontal surface entirely membranous. Four short spines are spaced around the operculum and another 10 pairs of equally short, thin, and straight or only slightly curved spines distributed along the margins of the frontal membrane. Each autozooid budding its successor from the basal wall, in the proximal third of the broad portion. Between this budding point and the filiform portion is a pair of short, tubular processes; on rare occasions one of these may develop as a zooid bud, allowing colony to branch, but most autozooids uniserially arranged.
Tentacles clear, 19–21; lophophores bellshaped, radially symmetrical.
DISTRIBUTION: Probably common in coastal habitats throughout the northeast Atlantic and the Mediterranean, on a wide variety of organic and inorganic substrata, but its inconspicuous colonies are easily overlooked.
Beania mirabilis has been described also from IndoPacific localities, but these records require reexamination.
MEASUREMENTS (CYSTID): ZL (distal) 677 ± 45 µm, 620–780 (2, 13), ZW (distal) 280 (1, 1). (POLYPIDE): IH 295 ± 59 µm, 229– 343 (2, 3), LD 642 ± 130, 491–806 (2, 4), TL 532 ± 128, 395–689 (2, 9).
FAMILY EPISTOMIIDAE GREGORY, 1893 View in CoL
GENUS SYNNOTUM PIEPER, 1881 View in CoL
Synnotum aegyptiacum ( Audouin, 1826) View in CoL Figure 10E, F View Fig
Loricaria aegyptiaca Audouin, 1826: 243 ; Savigny, 1809: pl.13, figs 4(1)–4(5).
Synnotum aegyptiacum: Prenant and Bobin, 1966: 461 View in CoL . Zabala and Maluquer, 1988: 100.
DESCRIPTION (AMNH 909; CMRR 2212): Colony erect, branching, jointed, to 7.5 mm in present material. Autozooids slender, fusiform, narrowest proximally, broadest distally; arranged in backtoback pairs, each separated from preceding pair by a slender, tubular chitinous node. Frontal surface entirely membranous, with a terminal operculum slightly acute to frontal plane. Squat, sessile avicularia present on distobasal surfaces of some autozooids, others bearing larger, pedunculate, bird’s head type with short, hooked mandible. Embryos brooded in distinctive globular, dimorphic zooids (not found in present material). Each autozooid of a pair giving rise to a single bud distobasally, which traverses the basal wall junction of the succeeding autozooid pair as a slender tube, then expands distal to them to form broad distal portion of new autozooid. Autozooid thus consisting of a proximal tube comprising twothirds its total length and a fusiform distal portion bearing the frontal membrane. The node between each autozooid pair, consequently, consisting of four closely juxtaposed tubes.
DISTRIBUTION: Widespread and common throughout the Mediterranean, in shallow coastal waters, and recorded from warm temperate and tropical seas worldwide.
MEASUREMENTS (SKELETAL): ZL 500 ± 41 µm, 430–577 (2, 21).
FAMILY CANDIDAE D’ORBIGNY, 1851
GENUS SCRUPOCELLARIA LEVINSEN, 1909 View in CoL
Scrupocellaria delilii ( Audouin, 1826) View in CoL Figure 10G–I View Fig
Crisia delilii Audouin, 1826: 242 . Savigny, 1809: pl. 12, fig. 3.
Scrupocellaria macandrei: Heller, 1867: 87 .
Scrupocellaria delilii: Gautier, 1962: 86 View in CoL . Prenant and Bobin, 1966: 435.
DESCRIPTION (AMNH 910; CMRR 2213): Colonies erect, branching dichotomously, to 20 mm high in present material. Branching frequency variable, at 2mm intervals in proximal regions of colony, to 4 mm or more towards distal portion; each ramus originating as a bipartite, tubular chitinous joint. Branches flat, consisting of two alternating longitudinal autozooid series; opesiae of adjacent autozooids overlapping for about onefifth of total length. Autozooids ca 0.5 X 0.25 mm; opesia oval, comprising more than half total frontal length, bordered by finely granular cryptocystal rim. Two spines present, one on each distal corner of opesia, erect, slender and curved, converging medially above operculum. A small scutum present proximal to inner distal spine, with a slender stalk curving over frontal membrane; oval, or slightly crescentic, consisting largely of a proximal lobe, with only a minimal lobe distal to stalk. Each autozooid with a prominent lateral avicularium, its cystid half as long as opesia, rostrum 0.15 mm long, sharply hooked distally and projecting distinctly from branch margin. Small frontal avicularia present on some autozooids, proximal to opesia, with short, triangular rostrum perpendicular to branch axis. Ovicell short, broad, tilted towards branch axis, imperforate. A small vibraculum on abfrontal surface of each autozooid, with setal groove straight and extending halfway to middle of the autozooid; two such vibracula in axil of each dichotomy.
DISTRIBUTION: Scrupocellaria delilii has been reported from the Atlantic coasts of Spain, south to Madeira, and from numerous localities in the Mediterranean. It has also been recorded from the Red Sea, the Indian Ocean, Australia and the West Pacific, but this wider Distribution possibly encompasses
the geographical ranges of more than one species.
MEASUREMENTS (SKELETAL): DO 323 ± 47 µm, 245–413 (2. 20), OpL 217 ± 21, 178– 243 (2, 20), OpW 137 ± 15, 107–161 (2, 20), ZL 425 ± 31, 383–503 (2, 20), ZW 219 ± 16, 188–251 (2, 20).
Scrupocellaria scrupea Busk, 1851 View in CoL Figure 11A–E View Fig
Scrupocellaria scrupea Busk, 1851: 83 View in CoL . Hayward and Ryland, 1998: 274.
DESCRIPTION (AMNH 911–913; CMRR 2214): Colonies to 18 mm in present material, dichotomously branching at frequent intervals, giving dense, bushy form. Opesia occupying half or less of total autozooid length; each autozooid with three spines on outer distal angle, two on the inner. Scutum arises close to base of proximalmost inner spine; lobe reniform, not pointed distally, covering almost half the area of the opesia. Prominent lateral avicularium present on each autozooid, with hooked rostrum, but no frontal avicularia. Basal vibraculum present at proximal end of each autozooid, aligned distoproximally, and two at axil of each dichotomy. Ovicell broader than long, inclined towards median longitudinal axis of the branch; lightly calcified, with a single, irregular foramen close to aperture.
Tentacles clear, 11; lophophores bellshaped, radially symmetrical.
DISTRIBUTION: In shallow coastal habitats throughout the Mediterranean, and north to the southwest British Isles.
MEASUREMENTS (SKELETAL): DO 319 ± 66 µm, 246–459 (2, 20), OpL 166 ± 16, 137– 195 (2, 20), OpW 109 ± 9, 95–129 (2, 20), ZL 378 ± 31, 326–435 (2, 20), ZW 201 ± 17, 177–233 (2, 20). (POLYPIDE): IH 13 ± 23 µm, 0–40 (1, 4), LD 273 ± 31, 240–300 (1, 4), TL 327 ± 23, 300–340 (1, 4).
Scrupocellaria scruposa ( Linnaeus, 1758) View in CoL Figure 11F–J View Fig
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Anguinaria truncata Landsborough, 1852: 288
| HAYWARD, PETER J. & McKINNEY, FRANK K. 2002 |
Scruparia ambigua : Hayward and Ryland, 1998: 108
| Hayward, P. J. & J. S. Ryland 1998: 108 |
Ellisina gautieri Fernández Pulpeiro and Reverter Gil, 1993: 98
| Hayward, P. J. & J. S. Ryland 1998: 192 |
| Fernandez Pulpeiro, E. & O. Reverter Gil 1993: 98 |
Copidozoum tenuirostre : Prenant and Bobin, 1966: 257
| Hayward, P. J. & J. S. Ryland 1998: 180 |
| Prenant, M. & G. Bobin 1966: 257 |
Terminoflustra tenella :
| Prenant, M. & G. Bobin 1966: 191 |
Synnotum aegyptiacum :
| Zabala, M. & P. Maluquer 1988: 100 |
| Prenant, M. & G. Bobin 1966: 461 |
Chartella tenella : Gautier, 1962: 48
| Gautier, Y. V. 1962: 48 |
Beania hirtissima : Gautier, 1962: 95
| Prenant, M. & G. Bobin 1966: 557 |
| Gautier, Y. V. 1962: 95 |
Beania hirtissima
| Gautier, Y. V. 1962: 96 |
Beania robusta : Gautier, 1962: 99
| Zabala, M. & P. Maluquer 1988: 101 |
| Gautier, Y. V. 1962: 99 |
Beania magellanica : Gautier, 1962: 97
| Prenant, M. & G. Bobin 1966: 555 |
| Gautier, Y. V. 1962: 97 |
Scrupocellaria delilii : Gautier, 1962: 86
| Prenant, M. & G. Bobin 1966: 435 |
| Gautier, Y. V. 1962: 86 |
Bugula fulva
| Hayward, P. J. & J. S. Ryland 1998: 218 |
| Ryland, J. S. 1960: 86 |
Flustra tenella
| Hincks, T. 1887: 313 |
Diachoris hirtissima
| Hincks, T. 1886: 263 |
Diachoris hirtissima
| Hincks, T. 1881: 133 |
Membranipora tenuirostre
| Hincks, T. 1880: 70 |
Diachoris hirtissima
| Heller, C. 1867: 94 |
Scrupocellaria macandrei :
| Heller, C. 1867: 87 |
Anguinaria truncata
| Gautier, Y. V. 1962: 29 |
| Landsborough, D. 1852: 288 |
Diachoris magellanica
| Busk, G. 1852: 54 |
Scrupocellaria scrupea
| Hayward, P. J. & J. S. Ryland 1998: 274 |
| Busk, G. 1851: 83 |
Beania mirabilis
| Hayward, P. J. & J. S. Ryland 1998: 244 |
| Alvarez, J. A. 1990: 26 |
| Johnston, G. 1840: 272 |
Flustra dumerilii
| Audouin, J. V. 1826: 240 |
Loricaria aegyptiaca
| Audouin, J. V. 1826: 243 |
Crisia delilii
| Audouin, J. V. 1826: 242 |