
Eschara fascialis Pallas, 1766: 42
|
publication ID |
https://doi.org/10.1206/0003-0090(2002)270<0001:NABFTV>2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/03D1878C-197E-FFE0-FD52-C69BFE55C4EE |
|
treatment provided by |
Felipe |
|
scientific name |
Eschara fascialis Pallas, 1766: 42 |
| status |
|
Eschara fascialis Pallas, 1766: 42 . Millepora foliacea Ellis and Solander, 1786: 133 . Eschara foliacea: Busk, 1854: 89 . Lepralia foliacea: Hincks, 1880: 300 .
Hippodiplosia foliacea: Canu and Bassler, 1930: 50 .
Pentapora fascialis: Zabala, 1986: 401 View in CoL . Hayward and Ryland, 1999: 200.
DESCRIPTION (AMNH 945–947; CMRR
2234): Colonies red, erect, rigid, heavily cal
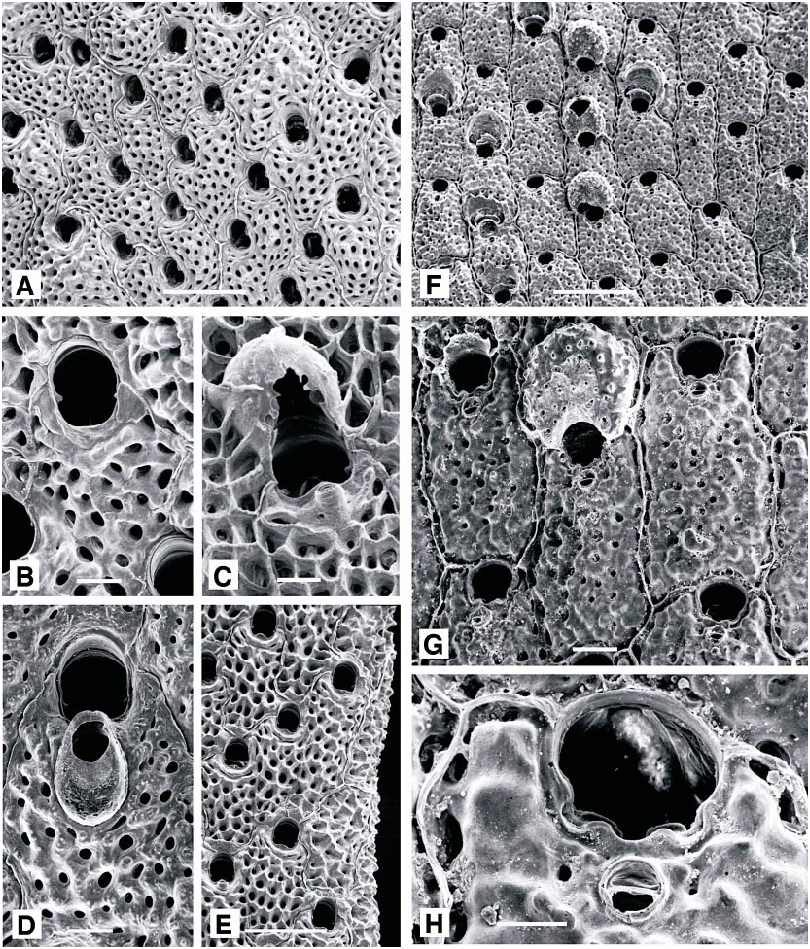
cified; consisting of bilaminar plates, dividing and anastomosing at irregular intervals, forming a threedimensional coralliform structure. Deep orange to redorange when living. Autozooids elongate, oval to hexagonal; distinct when newly budded, each separated by thick, raised ridges. Primary orifice narrowly bellshaped, longer than wide, with straight or slightly convex proximal margin between small, triangular, downcurved condyles. No oral spines. A small suboral avicularium sporadically present, missing from many autozooids; rostrum short, oval, slightly acute to frontal plane, proximally directed; occasionally replaced by a dimorphic avicularium with slightly spatulate rostrum, 0.2 mm long, normal to frontal plane, proximally directed. Frontal shield convex, evenly perforated; thickening rapidly from earliest ontogeny, with ridges developing between the pores, fusing to form a reticulate shield which continues to thicken through later on togeny, when individual autozooid boundaries become obscured. Ovicell elongate oval, with scattered frontal pores, visible only in early ontogeny, soon obscured by thickening calcification.
Tentacles light orange, 17–19; lophophores bellshaped, radially symmetrical away from colony edge, obliquely truncate along colony margins and locally (at chimneys?) on colony surface.
REMARKS: The colony habit is the narrow fronded, open form characteristic of Mediterranean populations. This is the largest and most conspicuous calcified bryozoan in the Adriatic, with colonies up to 0.5 m high and 0.3 m in diameter in the vicinity of Sv. Ivan Island before the anoxic events of the late 1980s. Colonies established after the anoxic event had grown to exceed 15 cm in diameter by summer 1998, although reestablishment of large colonies may require many more years due to exponential decrease in growth rate with increased size of colonies ( Cocito et al., 1998a). Recovery of P. fascialis from the northern Adriatic anoxic events of the late 1980s has been much slower than 3.5year recovery of the species at a shallower site in the Ligurian Sea following a severe storm in the early 1990s ( Cocito et al., 1998b). Where little disturbed by humans, P. fascialis normally grows in exposed positions; large colonies offshore of Rovinj commonly are found attached to gorgonians or flexible bryozoans, which reduces wave and current fragmentation. In other areas of the Mediterranean, P. fascialis grows in exposed positions on rock in depths as shallow as 11 m, although single severe storms can eliminate such shallow populations ( Cocito et al., 1998b). In areas frequented by divers, colonies are less frequent, smaller, and more cryptic ( Sala et al., 1996).
OCCURRENCE: With the exception of two localities (18, 19) approximately 20 m deep where colony fragments were dredged, Pentapora fascialis was found at 29 m and great er depths (summer thermocline is at about 29 m depth; Marinkovic´, 1959), growing on rock walls or more commonly on cobbles or on gorgonian stalks. Large colonies, over 30X20X 20 cm, were especially common on gorgonian stalks attached to cobbles and boulders on the sand floor northeast of Pelago Island at 30 m and greater depth.
DISTRIBUTION: Found from the Hebrides and the western coasts of Britain and Ireland southwards, and throughout the Mediterranean. Characteristic of currentswept coarse grounds.
MEASUREMENTS (SKELETAL): AL(normal) 93 ± 19 µm, 63–132 (2, 20), AW(normal) 71 ± 15, 43–101 (2, 20), AL(spatulate) 335 ± 29, 287–372 (1, 6), AW(spatulate) 188 ± 15, 163–206 (1, 6), DO 532 ± 102, 383–757 (2, 20), OL 197 ± 26, 164–247 (2, 20), OW 165 ± 20, 132–199 (2, 20), OvL 363 ± 20, 320–383 (1, 10), OvW 315 ± 17, 294–345 (1, 10), ZL 815 ± 70, 658–966 (2, 20), ZW 451 ± 40, 383–539 (2, 20). (POLYPIDE): IH 152 ± 95 µm, 60–440 (3, 19), LD 735 ± 121, 540–920 (3, 25), MDMn 28 ± 5, 20– 35 (2, 11), MDMx 34 ± 7, 20–40 (2, 11), TLMn 639 ± 135, 380–920 (3, 23), TLMx 694 ± 116, 400–920 (3, 24).
GENUS SCHIZOMAVELLA CANU AND BASSLER, 1917
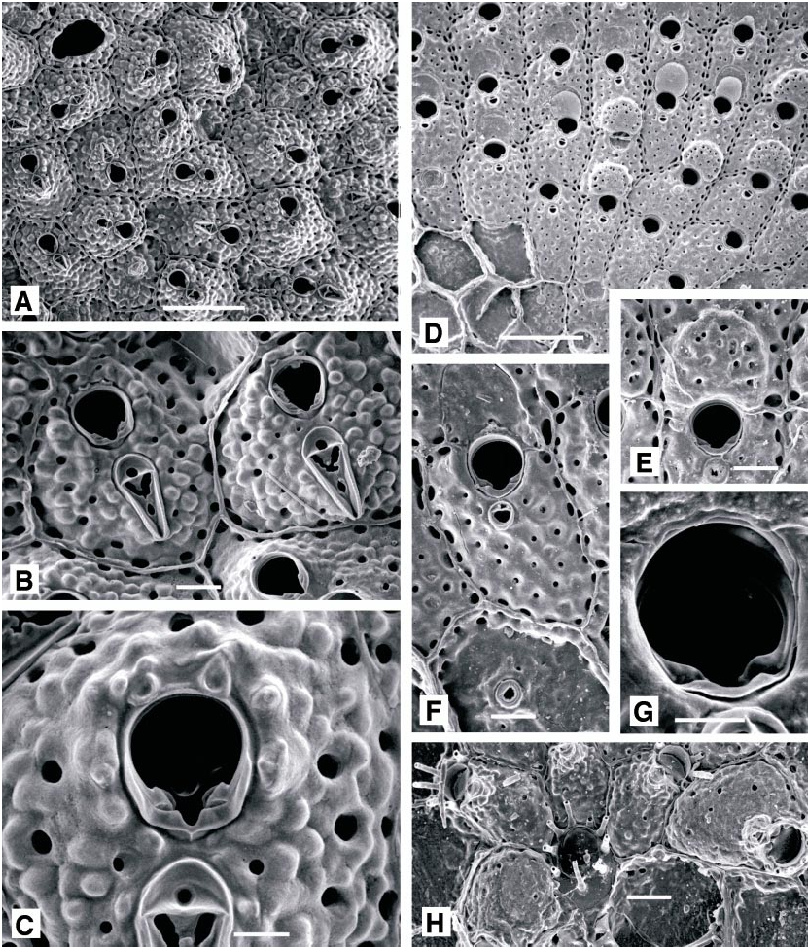
Schizomavella asymetrica ( Calvet, 1927) View in CoL Figure 25F–H View Fig
Schizoporella auriculata var. asymetrica Calvet, 1927: 20 . Gautier, 1962: 134.
Calyptotheca triarmata Hayward, 1974: 381 , fig. 5b.
Schizomavella cuspidata View in CoL (in part): ReverterGil and FernándezPulpeiro, 1995: 265 (fig. 4G only). Hayward and Ryland, 1999: fig. 131D only.
DESCRIPTION (AMNH 948): Colonies encrusting sheets, multiserial, unilaminar or with frontally budded patches. Autozooids rectangular to irregularly polygonal, rather flat, separated by distinct, raised sutures. Primary orifice wider than long, its greatest width at midlength; proximal border with a shallow, Ushaped median sinus, occupying onefifth of its width, accentuated by raised shoulders and broad, deep condyles which extend from the edges of the sinus, almost to lateral rim of orifice. Two or three indistinct distal oral spine bases visible in some autozooids, but most apparently lack spines entirely. Frontal shield thick, nodular, perforat ed by large, wellspaced pores, the marginal pores especially conspicuous. Avicularium median suboral, on a small prominence, close to proximal orifice rim but distinct from it; monomorphic, the rostrum parallel to frontal plane, proximally directed, small, broadly oval, with a complete crossbar but no palate. Ovicell distinctive: hyperstomial, recumbent on distally succeeding autozooid but projecting well above frontal plane; broader than long, the frontal surface pierced by as many as 20 small pores, developing a spiky, distal ooecial cover.
REMARKS: Elevation of Calvet’s (1927) variety asymetrica to full species rank is prompted by ReverterGil and Fernández Pulpeiro (1995), who published a micrograph showing the primary orifice in a specimen which they stated to be identical to material labeled ‘‘ var. asymetrica ’’ in Calvet’s hand. It is unquestionably the same species as that described and figured here. Reverter Gil and Fernández Pulpeiro (1995) considered var. asymetrica to fall within the range of variation displayed by S. cuspidata ( Hincks, 1880) as recently redefined (Hayward and Thorpe, 1995). However, the primary orifice of that species differs subtly in shape from that of var. asymetrica , being more rounded laterally, and the avicularia are dissimilar in the two species. In S. cuspidata the avicularium is always elongate oval and is polymorphic; the proximal rim of the rostrum is continuous with the proximal rim of the primary orifice; finally, in all avicularia there is a substantial palate with a characteristic trifoliate foramen. In S. asymetrica the avicularium is monomorphic, shortly oval and lacks a palate; further, it is distant from, and quite unattached to, the rim of the primary orifice. The frontal shield of S. cuspidata has fewer, smaller pores than that of S. asymetrica , and the ovicell is far less prominent, with fewer, larger pores, while the projecting ovicell of S. asymetrica bears up to 20 small pores frontally.
Calvet (1927) characterized var. asymetrica especially by the suboral avicularium, which he noted to be umbonate and laterally displaced with reference to the sinus. Gautier (1962) noted that in specimens from the western Mediterranean, the avicularium was either symmetrically or asymmetrically developed. In the present material the avicularium is consistently median suboral in position, but in specimens of what is certainly the same species from Chios, Aegean Sea, some autozooids display asymmetrical development of the avicularium, while in others a median avicularium may be accompanied by one or two lateral suboral avicularia. These specimens were described as Calyptotheca triarmata by Hayward (1974), and reexamination of the holotype specimen (NHM 1973.4.4.4) shows C. triarmata to be a junior subjective synonym of Schizomavella asymetrica ( Calvet, 1927) .
DISTRIBUTION: The species was first described from the vicinity of Monaco ( Calvet, 1927); Gautier (1962) recorded specimens from the same region and also from the Gulf of Marseille and Tunisia. The specimen illustrated by Hayward and Ryland (1999, fig. 131D) is a Norman collection specimen (NHM 1911.10.1.1538) from an unknown British locality.
MEASUREMENTS (SKELETAL): DO 378 ± 46 µm, 330–445 (1, 10), OL 88 ± 5, 82–97 (1, 10), OW 98 ± 5, 86–104 (1, 10), OvL 278 ± 17, 250–306 (1, 10), OvW 280 ± 28, 240–320 (1, 10), SL 11 ± 2, 9–15 (1, 10), SW 20 ± 2, 18–26 (1, 10), ZL 511 ± 27, 472–548 (1, 10), ZW 359 ± 20, 324–382 (1, 10).
Schizomavella cornuta ( Heller, 1867) View in CoL Figure 26A–D View Fig
Lepralia cornuta Heller, 1867: 110 .
Schizoporella auriculata var. cuspidata Hincks, 1880: 261 .
Schizomavella cuspidata: Hayward and Ryland, 1999: 286 View in CoL , fig. 131A–C (not 131D).
NEOTYPE (chosen here): UIIZ 270.
DESCRIPTION (AMNH 895, 949; CMRR 2235): Colonies creamcolored, encrusting, multiserial, unilaminar to multilaminar, often developing thick nodular formations. Autozooids rectangular in newly grown, unilaminar colonies, usually irregularly polygonal in multilaminar colonies developed through frontal budding; individual boundaries generally distinct, as narrow, raised ridges. Primary orifice characteristic: wider than long, with greatest width at midlength; proximal border with a short, symmetrical, Ushaped sinus defined by short lobes of the proximal orifice rim, proximal corners of orifice deeply notched; condyles short, broad and thick, extending from, and emphasizing, the sinus into the proximal corners of the orifice. Two to six distal oral spines present in early ontogeny, commonly obscured during later ontogeny. Ancestrula small, tatiform, opesia surrounded by nine marginal spines. Avicularium suboral, proximally directed, with rostrum almost parallel to frontal plane; dimorphic: short, oval, ca. 0.06 mm long, or ca. 0.2 mm long, narrowly spoonshaped, with slightly concave sides and gently expanded distal end. In both types of avicularia, proximal rim of rostrum continuous with outer rim of sinus, and a thick cross bar with a stout columella divides proximal portion of narrow palatal foramen to form a distinctive Y shape. Ovicell broader than long, flattened frontally, with few (ca. 5) large, irregular pores immediately distal to the arched aperture; developing a nodular ooecial cover in later ontogeny.
Embryo color medium orange to dark, brickred.
REMARKS: As Schizomavella cuspidata , this species has only recently been redescribed and distinguished from S. auriculata (Hassall) , with which it was for long confused (Hayward and Thorpe, 1995). However, as recognized by Friedl (1917: 272), part of Heller’s material of Lepralia cornuta in the University of Innsbruck collections has affinities with Schizomavella auriculata (referred to as Smittina auriculata by Heller). We designate UIIZ specimen 270, illustrated here as fig. 26A and 26B, as neotype of Lepralia cornuta because it may not have been in Heller’s possession when he named the species. If eventually discovered to be a specimen on which Heller originally described Lepralia cornuta , it should be redesignated as lectotype. UIIZ 270 is conspecific with S. cuspidata , and Lepralia cornuta is senior objective synonym of the two.
DISTRIBUTION: It is common on northwest European coasts, although its geographical and ecological Distribution is still incompletely known. In the Mediterranean its occurrence is even more imprecisely described through confusion with numerous other species of Schizomavella . The ovicell figured by Hayward and Ryland (1999, fig. 131D) is not that of S. cuspidata , but rather S. asymetrica , here redescribed for the first time.
MEASUREMENTS (SKELETAL): AL(normal) 63 ± 9 µm, 48–81 (2, 20), AW(normal) 68 ± 8, 54–84 (2, 20), AL(spatulate) 149 ± 16, 131–175 (1, 6), AW(spatulate) 65 ± 9, 55– 76 (1, 6), DO 409 ± 66, 317–526 (2, 20), OL 86 ± 6, 75–98 (2, 20), OW 102 ± 4, 94–104 (2, 20), SL 20 ± 2, 16–24 (2, 20), SW 27 ± 4, 21–34 (2, 20), ZL 458 ± 39, 383–510 (2, 20), ZW 347 ± 52, 244–460 (2, 20).
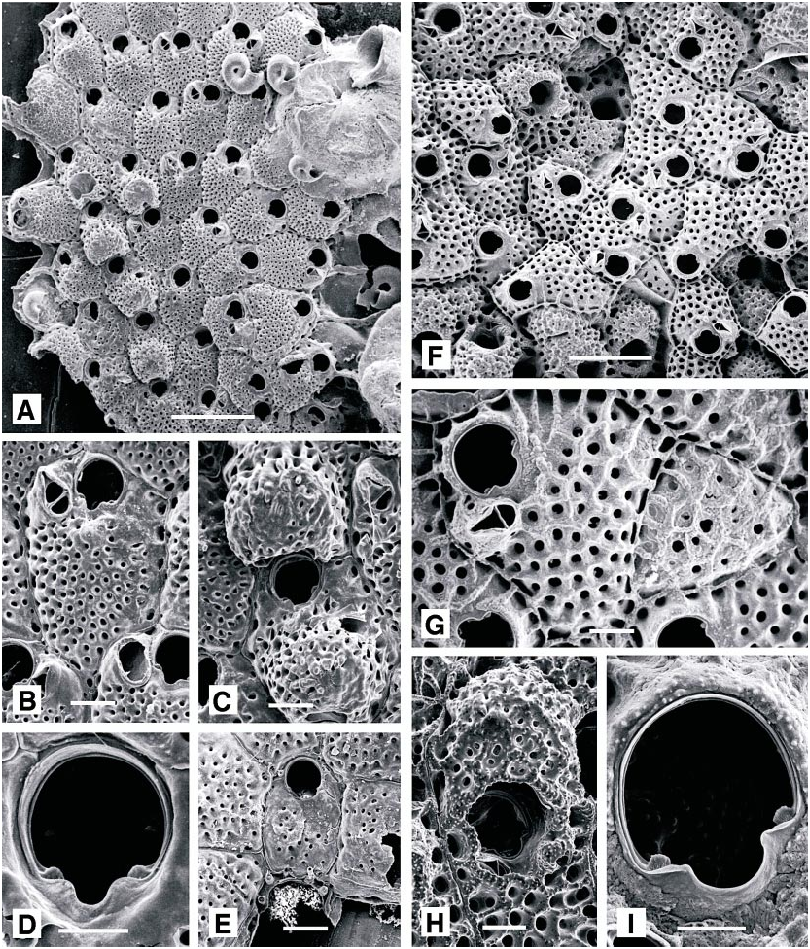
Schizomavella linearis ( Hassall, 1841) View in CoL Figure 26E–G View Fig
Lepralia linearis Hassall, 1841: 368 .
Schizomavella linearis: Gautier, 1962: 140 View in CoL . Zabala, 1986: 473. Zabala and Maluquer, 1988: 132. Hayward and Thorpe, 1995: 671.
DESCRIPTION (AMNH 950; CMRR 2236): Colonies encrusting, multiserial, unilaminar to multilaminar sheets. Autozooids quadran
gular to polygonal, rather flat, vertical walls distinct; frontal shield nodular, with conspicuous marginal perforations, and few, large pores medially. Primary orifice as wide as long, broadest at midlength; sinus short, Ushaped, occupying about half width of proximal border; condyles short, broad and rounded. Two or three slender distal oral spines present. Avicularia single or paired, lateroproximal to orifice; rostrum narrowly acuminate, typically directed medially or proximomedially, occasionally distally. Large adventitious avicularia sporadic, characteristic: cystid domed, occupying entire frontal shield of bearing autozooid, mandible narrowly crescentic, 0.25 mm wide. Ovicells broader than long, flattened frontally, with numerous irregular pores and an umbonate ooecial cover.
DISTRIBUTION: This species is common on north European coasts. It has been recorded previously in the Mediterranean but is perhaps far less frequent on southern European coasts. It was the least common of the described species of Schizomavella found at Rovinj.
MEASUREMENTS (SKELETAL): AL(normal) 107 ± 9 µm, 88–121 (2, 21), AW(normal) 88 ± 10, 70–105 (2, 21), DO 412 ± 78, 315– 576 (2, 20), OL 108 ± 10, 90–122 (2, 20), OW 91 ± 8, 81–106 (2, 20), OvL 256 ± 22, 236–301 (1, 10), OvW 297 ± 30, 282–329 (1, 10), SL 24 ± 4, 19–29 (2, 20), SW 26 ± 7, 16–38 (2, 20), ZL 491 ± 47, 426–586 (2, 20), ZW 333 ± 50, 265–461 (2, 20).
Schizomavella mamillata ( Hincks, 1880) View in CoL Figure 27A–C View Fig
Schizoporella linearis var. mamillata Hincks, 1880: 248 View in CoL .
Schizomavella mamillata: Gautier, 1958: 199 View in CoL . Zabala, 1986: 475. Zabala and Maluquer, 1988: 132.
Schizomavella linearis var. mamillata: Gautier, 1962: 143 View in CoL . Harmelin, 1969 a: 1201.
DESCRIPTION (AMNH 951, 952; CMRR 2237): Colonies orange, encrusting, multiserial, multilaminar, developing a massive mamillate form. Autozooids rectangular to irregularly polygonal, in regular series at unilaminar growing edges, randomly orientated in areas of vigorous frontal budding; boundaries always distinct, marked by thin raised sulci. Primary orifice longer than wide, tapered proximally, midproximal border with a short, symmetrical rounded notch; thick, faceted condyles on each side of notch define a deep, Vshaped sinus. Oral spines absent to four, most commonly two. Peristome developed as a thin, inconspicuous rim, most pronounced, and slightly flared, on each side of sinus. Frontal shield convex, nodular and vitreous, evenly perforated. Avicularium diagnostic: median suboral, monomorphic, directed proximally, slightly acute to frontal plane; rostrum elongate triangular, its tip downcurved and finely denticulate; crossbar complete, with median columella, palate with trifoliate foramen. Ovicell hyperstomial, prominent, about as wide as long, with 10– 15 large pores frontally.
Tentacles light orange, 14–15; lophophores bellshaped, radially symmetrical between chimneys and obliquely truncate adjacent to chimneys.
REMARKS: We here designate as lectotype BMNH specimen 1899.5.1.1059, Hincks Collection, from Algiers.
DISTRIBUTION: Apparently widespread and common throughout the Mediterranean, on shallow, coarse detrital grounds.
MEASUREMENTS (SKELETAL): AL 191 ± 26 µm, 153–247 (2, 210), AW 127 ± 17, 104– 151 (2, 21), DO 520 ± 90, 351–664 (2, 20), OL 106 ± 11, 94–141 (2, 20), OW 100 ± 8, 88–116 (2, 20), SL 28 ± 6, 19–36 (2, 20), SW 32 ± 4, 25–42 (2, 20), ZL 690 ± 70, 563–822 (2, 20), ZW 398 ± 45, 351–502 (2, 20). (POLYPIDE): IH 110 ± 37 µm, 60–180 (1, 7), LDMn 478 ± 65, 420–660 (1, 12), LDMx 500 ± 68, 420–660 (1, 12), MD 26 ± 2, 25–30 (1, 4), TLMn 514 ± 38, 470– 580 (1, 8), TLMx 540 ± 66, 480–680 (1, 8).
Schizomavella rudis (Manzoni, 1869) Figure 27D–H View Fig
Lepralia rudis Manzoni, 1869a: 18 .
Schizomavella rudis: Gautier, 1962: 146 . Zabala, 1986: 478. Zabala and Maluquer, 1988: 132. ReverterGil and FernándezPulpeiro, 1995: 261.
DESCRIPTION (AMNH 932, 953, 954 CMRR 2238): Colonies creamcolored, orange, or red; multiserial, encrusting, unilaminar. Autozooidal frontal wall uniformly po rous and with small areolae, lacking suboral umbo; autozooidal orifice roughly equidimensional, lacking distal spines and orificial peristome, proximal sinus relatively wide and defined by essentially smooth condyles. Suboral, small, oval, laterally or proximally orientated adventitious avicularium present on some autozooids. Ovicells prominent, emergent, perforated by pores distributed in a semilunar region partially encircling a proximal, porefree region adjacent to the orifice.
Tentacles 13–15; lophophores bellshaped, radially symmetrical midway between chimneys to strongly obliquely truncate (some campylonemidan) at chimneys and colony margins; adjacent lophophores overlapped up to at least 50 µm.
REMARKS: ReverterGil and Fernández Pulpeiro (1995) characterized S. rudis as usually having pores on the ovicell confined to a central, essentially circular area, but occasionally having them distributed in a semilunar region partially encircling a proximal, porefree region adjacent to the orifice. All specimens of S. rudis examined from the vicinity of Rovinj have the semilunar distribution of pores.
OCCURRENCE: Encrusts diverse bivalve substrata.
DISTRIBUTION: Gautier (1962) recorded this species from numerous localities in the western Mediterranean, from 20–80 m depth, most often encrusting biogenic carbonates.
MEASUREMENTS (SKELETAL): AL 57 ± 11 µm, 50–70 (3, 15), AW 52 ± 13, 40–90 (3, 15), DO 377 ± 64, 270–480 (3, 30), OL 101 ± 9, 80–110 (3, 15), OW 100 ± 10, 80–120 (3, 15), ZL 530 ± 62, 440–620 (3, 15), ZW 360 ± 55, 300–480 (3, 15). (POLYPIDE): IH 165 ± 74 µm, 100–310 (2, 15), LDMn 556 ± 82, 440–720 (2, 12), LDMx 608 ± 113, 460–780 (2, 18), MD 25.7 ± 2.6, 25–30 (2, 16), TLMn 462 ± 98, 300–600 (2, 13), TLMx 649 ± 134, 400–840 (2, 16).
Schizomavella subsolana , new species Figure 28A–D View Fig
HOLOTYPE: AMNH 955.
PARATYPES: AMNH 956, 901; CMRR 2239–2240.
DIAGNOSIS: Primary orifice nearly equidi
mensional, with shallow sinus; three to five distal oral spines; single small suboral avicularium with elongate triangular rostrum, steeply inclined to plane of orifice.
ETYMOLOGY: The species name is from the Latin sub (under) and sol (sun) in recognition of its discovery along a coast that is a popular summer holiday destination.
DESCRIPTION: Colonies creamcolored, small, rounded, multiserial, unilaminar patches. Autozooids in regular, radiating lines, oval to hexagonal, separated by deep grooves. Frontal shield steeply convex, distinctly umbonate distally, umbo incorporating median, suboral avicularium; rugose, evenly perforated by small, round pores. Primary orifice slightly wider than long, a shallow sinus occupying more than half its proximal width, between short, inconspicuous condyles. Three to five (commonly four) distal oral spines present; peristome developed as a low rim, extending, on each side, from proximalmost spine to avicularium, deepest proximally. Avicularium almost perpendicular to plane of orifice, rostrum elongate triangular. Ovicell slightly broader than long, flattened frontally, with 20–30 irregular pores; initially smooth, but developing a coarse ooecial cover, coextensive with the frontal shield of distally succeeding autozooid.
REMARKS: The taxonomy of north European species of Schizomavella was reviewed and revised by Hayward and Thorpe (1995), who redescribed and illustrated five species. Subsequently, new species have been described from both north European and Mediterranean localities ( ReverterGil and FernándezPulpeiro, 1995, 1997), and the taxonomy of some west Mediterranean species has also been clarified. However, the systematics of the genus in the Mediterranean as a whole remains in need of revision. This new species differs from all of those recently described or redescribed; its distinctive suite of characters includes the primary orifice, with an especially broad sinus, three to five oral spines, and the comparatively large, frontally directed avicularium. It does not seem to have been noted in any of the literature treating ‘‘varieties’’ of Mediterranean Schizomavella species (e.g., Gautier, 1962).
MEASUREMENTS (SKELETAL): AL 88 ± 11 µm, 74–115 (2, 20), AW 95 ± 12, 78–130 (2, 20), DO 366 ± 42, 274–433 (3, 30), OL 85 ± 6, 78–100 (2, 12), OW 88 ± 8, 78– 104 (2, 20), OvL 201 ± 27, 155–248 (3, 30), OvW 205 ± 15, 178–239 (3, 30), SL 18 ± 7, 9–34 (2, 12), SW 50 ± 6, 35–56 (2, 12), ZL 431 ± 30, 386–490 (3, 30), ZW 309 ± 36, 275–339 (3, 30).
FAMILY WATERSIPORIDAE VIGNEAUX, 1949 View in CoL GENUS WATERSIPORA NEVIANI, 1895 View in CoL Watersipora complanata ( Norman, 1864) View in CoL Figure 28E, F View Fig
Lepralia complanata Norman, 1864: 85 .
Watersipora complanata: Hayward and Ryland, 1999: 192 View in CoL .
DESCRIPTION (AMNH 957; CMRR 2241): Colonies brown, spreading multiserial, unilaminar sheets. Autozooids oval to hexagonal, flat or slightly convex, separated by distinct raised ridges. Primary orifice as wide as long, more or less bellshaped, with lateral rims curving inwards towards small, downcurved condyles just above the proximal bor der, which is straightedged but arched frontally. No spines. Orifice rim thickens in later ontogeny, forming pronounced distal and proximal lips. Frontal shield smooth, vitreous, glistening, densely perforated by small round pores, except for a small area immediately proximal to orifice. No avicularia . No ovicells.
DISTRIBUTION: Watersipora complanata is probably widely distributed in the Mediterranean. although it has been reported on relatively few occasions. Outside of the Mediterranean, it is known only from the Scilly Isles, southwest England, where it is common on the lower shore, encrusting a wide range of substrata.
MEASUREMENTS (SKELETAL): DO 471 ± 75 µm, 344–709 (3, 30), OL 121 ± 13, 100– 166 (3, 30), OW 171 ± 22, 135–211 (3, 30), ZL 692 ± 66, 588–813 (3, 30), ZW 390 ± 61, 277–503 (3, 30).
Watersipora subovoidea (d’Orbigny, 1852) Figure 29A, B View Fig
Cellepora ovoidea Audouin, 1826: 238 . Savigny,
1809: pl. 8, fig. 1.
Cellepora subovoidea d’Orbigny, 1852: 402 .
Lepralia cucullata Busk, 1854: 81 . Watersipora subovoidea: Gautier, 1962: 183 View in CoL . Zabala, 1986: 396.
DESCRIPTION (AMNH 958; CMRR 2242): Colonies spreading, multiserial, unilaminar, deep purple to black sheets to erect bilaminar plates (not seen in present material), often with zooidal boundaries and colonial organization difficult to recognize in living specimens. Autozooids with thick, deep purple to black epitheca, and deep black operculum. Autozooidal skeletons with densely and uniformly perforated cryptocystidean frontal shield. Primary orifice slightly wider than long, with robustly thickened rim and broad, Ushaped sinus occupying half of width of proximal edge between pointed, upturned condyles.
Tentacles bright red (carmine), 23–26; lophophores bellshaped, radially symmetrical, supported on long introverts.
REMARKS: All of our material formed jet black, unilaminar sheets encrusting rock, with the black color extending all the way to the growing edge, and with zooidal boundaries and opercula indistinguishable under the microscope when alive. Soule and Soule (1975) and Gordon (1989) discussed some of the tortuous synonymy of a group of confused watersiporids that includes ‘‘ W. subovoidea .’’ Gordon (1989) justified use of the epithet subtorquata in preference to subovoidea for the widespread fouling species that occurs in the Pacific ocean. Our specimens exhibit several differences from the characteristics of W. subtorquata as described and illustrated in Gordon (1989) and Gordon and Mawatari (1992). The New Zealand W. subtorquata has smaller (0.74–0.85 mm length), orange lophophores (very lightcolored in Gordon and Mawatari, 1992, Plate 1D) comprising 24 tentacles; colonies grade from black or dark gray in colony centers to dull orange peripherally, with broad orange margins; and the species is a common fouler in harbors and marinas, whereas specimens that we assign to W. subovoidea were found only on rock in clean water well away from harbors. W. subtorquata and W. subovoidea may be two names applied to a single, widely distributed and variable species, or more likely, as implied by Soule and Soule (1975) they refer to an inadequately discriminated species complex.
DISTRIBUTION: Throughout the Mediterranean, and for some unknown distance beyond, depending upon resolution of the taxonomy of the species or species complex.
MEASUREMENTS (SKELETAL): DO 682 ± 91 µm, 500–820 (2, 20), OL 239 ± 1 2, 219– 265 (2, 20), OW 249 ± 14, 216–266 (2, 20), SL 56 ± 11, 39–78 (2, 20), SW 127 ± 14, 96–148 (2, 20), ZL 998 ± 114, 772–1236 (2, 20), ZW 514 ± 65, 378–673 (2, 20). (POL YPIDE): IH 492 ± 81 µm, 340–700 (1, 17), LD 1261 ± 178, 1060–1620 (1, 12), MDMn 35 ± 6, 30–40 (1, 4), MDMx 42.5 ± 5, 40– 50 (1, 4), TL 1154 ± 129, 900–1400 (1, 16), TN 25 ± 1.3, 23–28 (1, 16).
SUPERFAMILY SCHIZOPORELLOIDEA JULLIEN, 1882 View in CoL
FAMILY SCHIZOPORELLIDAE JULLIEN, 1882 View in CoL
GENUS SCHIZOBRACHIELLA CANU AND BASSLER, 1920 View in CoL
Schizobrachiella sanguinea ( Norman, 1868) View in CoL Figure 29C–F View Fig
Hemeschara sanguinea Norman, 1868: 222 .
Schizoporella sanguinea: Hincks, 1880: 252 . Harmer, 1902: 303.
Schizobrachiella sanguinea: Canu and Bassler, 1930: 32 View in CoL . Gautier, 1962: 128. Zabala, 1986: 460. Zabala and Maluquer, 1988: 130. Hayward and Ryland, 1999: 222.
DESCRIPTION (AMNH 959, 960; CMRR 2243): Colonies with bright red cuticle, encrusting, multiserial, unilaminar to multilaminar, occasionally as erect, folded plates only partly attached to the substratum; deep red when living, dull brown when dead and dried. Autozooids rectangular to irregularly polygonal, flat or slightly convex, separated by thin, raised sutures. Primary orifice wider than long; proximal border with a short, Ushaped sinus occupying about onethird its width, defined by prominent shoulders which intersect on each side with broad, angular condyles, such that orifice appears proximally tridentate. No oral spines or peristome. Frontal shield densely perforated by large round pores, each bordered by a thickened, raised rim. Avicularia adventitious, monomorphic, sporadic: typically developed on autozooid margins, adjacent or distal to ori fice; small, <0.1 mm long, inconspicuous, rostrum acute or rounded, orientation variable. Ovicell recumbent on succeeding autozooid, domed and prominent, slightly wid er than long; densely perforated by small round pores and appearing coarsely tuberculate, closed by autozooid operculum.
Tentacles bright red to purple, 14–20 but with smaller range within individual colonies; lophophores bellshaped, radially symmetrical between chimneys, obliquely truncate adjacent to chimneys; adjacent lophophores overlapped up to 100 µm.
Embryos dark purple to black. Ancestrula and astogeny were described by Friedl (1925).
REMARKS: Many colonies are infested with a commensal stoloniferous hydrozoan ( Halocoryne epizoica Hadzi, 1917 , according to Gautier, 1962: 394). The relationship is apparently obligate for the hydrozoan ( Gautier, 1962) but not for the host species.
DISTRIBUTION: This distinctive species is widespread throughout the Mediterranean, encrusting organic carbonates in particular, and ranges north to the western approaches to the English Channel, and to southwest Cornwall, where it is rare. Harmelin et al. (1989) note the similarity between S. sanguinea , type species of Schizobrachiella Canu and Bassler, 1920 , and species of the warm temperate to tropical genus Calyptotheca Harmer, 1957 . Three species of Calyptotheca have been described from Mediterranean localities in recent years ( Hayward, 1974; Harmelin et al., 1989), although one, C. triarmata Hayward , is here shown to be a synonym of Schizomavella asymetrica (Calvet) .
MEASUREMENTS (SKELETAL): DO 514 ± 78 µm, 410–715 (2, 20), OL 162 ± 15, 135– 188 (2, 20), OW 182 ± 19, 153–209 (2, 20), OvL 318 ± 60, 236–392 (2, 6), OvW 364 ± 30, 327–400 (2, 6), SL 16 ± 3, 10–20 (2, 20), SW 31 ± 4, 25–37 (2, 20), ZL 474 ± 43, 386–548 (2, 20), ZW 670 ± 46, 598–759 (2, 20). (POLYPIDE): IH 108 ± 58 µm, 10– 260 (4, 38), LDMn 674 ± 113, 500–900 (4, 43), LDMx 705 ± 131, 500–1080 (4, 51), MD 38 ± 2, 30–40 (4, 20), TLMn499 ± 95, 380–780 (4, 30), TLMx 596 ± 139, 380– 1000 (4, 38).
GENUS SCHIZOPORELLA HINCKS, 1877 View in CoL ( nomen protectum)
Schizoporella Hincks, 1877 View in CoL , is apparently a junior subjective synonym of Multiporina d’Orbigny, 1852 , but the younger name is valid according to section 23.9.1 of the International Code of Zoological Nomenclature ( Ride et al., 1999: 28).
The genus Multiporina was established by d’Orbigny (1852: 445) for his new species Multiporina ostracites (d’Orbigny, 1852: 445) from Oligocene deposits of Montmartre, Paris, France as type species. d’Orbigny gave only a diagnosis with no illustration, and the diagnosis was considered subsequently to be insufficient for the genus to be recognizable, for example, Bassler (1935: 152, 1953: G235). The name Multiporina has not been used to our knowledge since 1899 (except to note that it should be suppressed: Canu, 1908; Buge, 1975; Gordon, 1989, or is unrecognizable: Bassler, 1935, 1953).
Schizoporella View in CoL was established by Hincks (1877: 527), with Lepralia unicornis Johnston in Wood (1844: 19) from the Pliocene Crag deposits of East Anglia as type species. Johnston (1847: 320, pl. 57, fig. 1), often is cited as the publication in which Lepralia unicornis was named. Recognition of the 1844 publication is important not only for the date and any implications of priority with respect to other names, but also because the 1847 publication describes living, not fossil material, which potentially is of importance in characterizing the species.
The genus name Schizoporella View in CoL has been used in many more than 25 works (probably thousands of works) published by at least 10 authors during the past 50 years. Species of Schizoporella View in CoL are prominent among marine fouling faunas (e.g., Geraci and Relini, 1970; Occhipinti Ambrogi, 1981; Brock, 1985), and in many instances they cover a high proportion of the substratum ( Sutherland, 1977, 1978). The genus is species rich, found widely in the Atlantic and Pacific Oceans, ranges from shallow equatorial waters (e.g., Cook, 1985) to the highest latitudes (e.g., Kluge, 1975), is diverse and abundant in temperate waters (e.g., Maturo, 1957; Gautier, 1962; Winston, 1982; Zabala, 1986; Zabala and Maluquer, 1988; Gordon, 1989; Hayward and Ryland, 1999) and has an abundant fossil record (e.g., Ziko, 1985; Spencer and Campbell, 1987; Poluzzi et al., 1988; Moissette and Saint Martin, 1995). Consequently it is among the most frequently identified and recorded bryozoan genera by marine biologists in general, not just by bryozoan specialists. Schizoporella View in CoL is noted as an exemplar in studies of benthic ecology (e.g., Buss, 1981; Lindberg and Stanton, 1989; Hurlbut, 1991; McKinney, 1992; Herrera and Jackson, 1996; Cocito et al., 2000), in comparative studies of feeding and related functions ( Winston, 1977, 1978; M. J. McKinney, 1997), in evolutionary research ( Schopf and Dutton, 1976), and among diverse other topics.
Canu (1908: 86) examined d’Orbigny’s holotype of Multiporina ostracites and placed it in synonymy with Schizoporella unicornis , giving preference to both the trivial name unicornis and the generic name Schizoporella . Buge (1975: 50, pl. 4, fig. 3) also examined, and illustrated, d’Orbigny’s holotype of Multiporina ostracites , agreeing with Canu in considering it to be synonymous with Schizoporella unicornis . Probably ostracites is not the same species as unicornis , inasmuch as cheilostome species seldom have a geological range sufficiently long to encompass both the Pliocene and Oligocene. Even if Multiporina ostracites and Schizoporella unicornis are not synonymous, they are closely related within the same genus. This interpretation is based on similarity of zooidal characters such as texture of perforations of the frontal wall; placement and shape of the orifice; and placement, shape, size, and orientation of the single adventitious avicularium per zooid. We here accept that Schizoporella is the valid name, and Multiporina should be suppressed.
Schizoporella unicornis (Johnston in Wood, 1844) Figure 30A–E View Fig



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Eschara fascialis Pallas, 1766: 42
| HAYWARD, PETER J. & McKINNEY, FRANK K. 2002 |
Schizomavella cuspidata : Hayward and Ryland, 1999: 286
| Hayward, P. J. & J. S. Ryland 1999: 286 |
Watersipora complanata : Hayward and Ryland, 1999: 192
| Hayward, P. J. & J. S. Ryland 1999: 192 |
Schizoporella
| Ride, W. D. L. & H. G. Cogger & C. Duuis & O. Kraus & A. Minelli & F. C. Thompson & P. K. Tubbs 1999: 28 |
Schizomavella cuspidata
| Reverter-Gil, O. & E. Fernandez-Pulpeiro 1995: 265 |
Pentapora fascialis : Zabala, 1986: 401
| Hayward, P. J. & J. S. Ryland 1999: 200 |
| Zabala, M. 1986: 401 |
Calyptotheca triarmata
| Hayward, P. J. 1974: 381 |
Schizomavella linearis :
| Zabala, M. & P. Maluquer 1988: 132 |
| Zabala, M. 1986: 473 |
| Gautier, Y. V. 1962: 140 |
Schizomavella linearis var. mamillata :
| Gautier, Y. V. 1962: 143 |
Schizomavella rudis : Gautier, 1962: 146
| Reverter-Gil, O. & E. Fernandez-Pulpeiro 1995: 261 |
| Zabala, M. & P. Maluquer 1988: 132 |
| Zabala, M. 1986: 478 |
| Gautier, Y. V. 1962: 146 |
Schizomavella mamillata : Gautier, 1958: 199
| Zabala, M. & P. Maluquer 1988: 132 |
| Zabala, M. 1986: 475 |
| Gautier, Y. V. 1958: 199 |
foliacea :
| Canu, F. & R. S. Bassler 1930: 50 |
Schizobrachiella sanguinea : Canu and Bassler, 1930: 32
| Hayward, P. J. & J. S. Ryland 1999: 222 |
| Zabala, M. & P. Maluquer 1988: 130 |
| Zabala, M. 1986: 460 |
| Gautier, Y. V. 1962: 128 |
| Canu, F. & R. S. Bassler 1930: 32 |
Schizoporella auriculata var. asymetrica
| Gautier, Y. V. 1962: 134 |
| Calvet, L. 1927: 20 |
Schizoporella auriculata var. cuspidata
| Hincks, T. 1880: 261 |
Schizoporella linearis var. mamillata
| Hincks, T. 1880: 248 |
Schizoporella sanguinea :
| Harmer, S. F. 1902: 303 |
| Hincks, T. 1880: 252 |
Lepralia rudis
| Manzoni, A. 1869: 18 |
Hemeschara sanguinea
| Norman, A. M. 1868: 222 |
Lepralia cornuta
| Heller, C. 1867: 110 |
Lepralia complanata
| Norman, A. M. 1864: 85 |
Lepralia cucullata
| Zabala, M. 1986: 396 |
| Gautier, Y. V. 1962: 183 |
| Busk, G. 1854: 81 |
Lepralia linearis
| Hassall, A. H. 1841: 368 |
Cellepora ovoidea
| Audouin, J. V. 1826: 238 |
Eschara fascialis
| Hincks, T. 1880: 300 |
| Busk, G. 1854: 89 |
| Ellis, J. & D. Solander 1786: 133 |
| Pallas, P. S. 1766: 42 |