
Acanthascus koltuni, Reiswig & Stone, 2013
|
publication ID |
https://doi.org/10.11646/zootaxa.3628.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:37D2D7F2-FA0C-40E9-B6D0-9C74EBB6C7F0 |
|
persistent identifier |
https://treatment.plazi.org/id/03D287B2-FF93-3606-9AD7-FCEE2A5FFD0B |
|
treatment provided by |
Felipe |
|
scientific name |
Acanthascus koltuni |
| status |
sp. nov. |
Acanthascus koltuni View in CoL n. sp.
( Figs. 21 View FIGURE 21 & 22 View FIGURE 22 , Table 11)
Synonymy. Acanthascus (Acanthascus) profundum ssp. nov. Stone et al., 2011: 26.
Material examined. Holotype: USNM# 1196559 About USNM , ROV ' Jason II' from RV ' Roger Revelle', dive J2099, 31 July 2004, Adak Canyon , 15.9 km SW of Cape Yakak, Adak Island , Aleutian Islands , Alaska, 51º28.091'N, 177º03.055'W, 2105 m, dry & ethanol. GoogleMaps
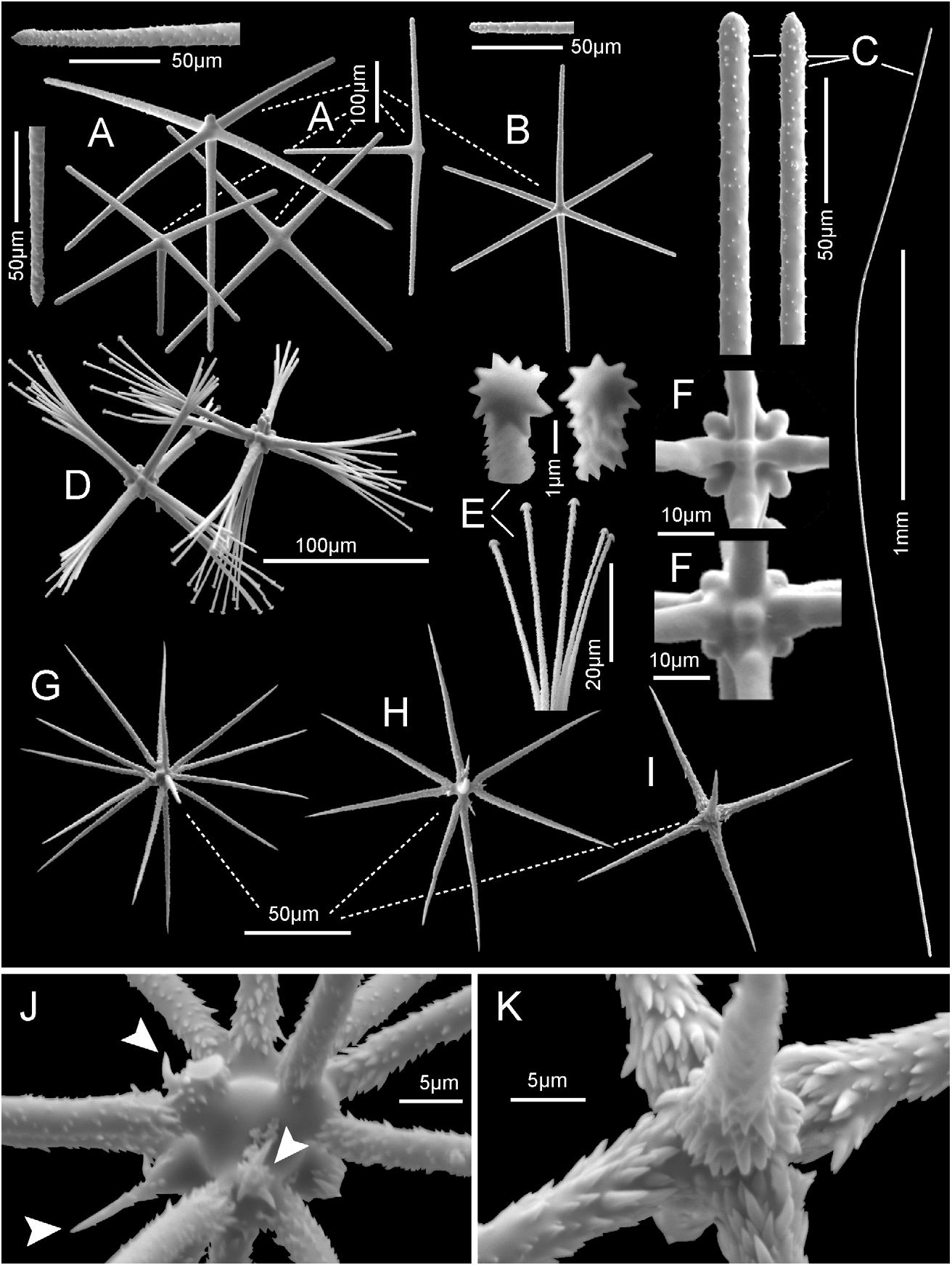
Description. The specimen encountered in situ ( Fig. 21A View FIGURE 21 ) was a white, thick-walled sac with very large external cones and relatively small circular osculum; before sampling, the specimen measured in situ with lasers was 30 cm tall and 38 cm in widest diameter. The deck photo of the collected upper 2/5ths ( Fig. 21B View FIGURE 21 ) shows it was 14.8 cm long by 9.6 cm wide and the large surface cones largely collapsed when removed from water; the basal part was not collected. The very delicate sponge was divided into three parts, one of which was transferred to 95% ethanol and the other two dried ( Fig. 21C View FIGURE 21 ). The wall is up to 2.2 cm thick. Prostalia are entirely lacking. The dermal surface retains indication of the collapsed large cones by small remaining cone tips from which radiate fine surface lines made by tissues associated with trains of hypodermal diactins that crisscross the entire surface. The hypodermal trains support a fine lattice of loose spicules ( Fig. 21D View FIGURE 21 ) covering an extensive subdermal space; inhalant canals leaving the subdermal space are 2–5 mm in diameter and are conspicuous through the dermal lattice. The atrial surface is extremely irregular, with large deep outpockets mostly 8–20 mm diameter extending into the wall; at a finer (mm) scale the surfaces of these outpockets receive small exhalant canals which open directly without a lattice covering ( Fig. 21E View FIGURE 21 ). The oscular margin is smooth and rather blunt, without marginalia. The main supporting skeleton is a network of loose interwoven unfused diactins, occurring singly or in bundles, oriented without order. The entire sponge is excessively cavernous and fragile; no spicule fusion is present in the part collected. The fresh specimen is white; when preserved in ethanol or dried it is light brown.
Megascleres: (spicule dimensions are given in Table 11). Hypodermalia are diactins; no hypodermal pentactins are present. Dermalia ( Fig. 22A View FIGURE 22 ) are mainly pentactins (90%) with a few hexactins (2%), irregular tetractins (5%), stauractins (2%), triactins (1%), and diactins (<1%); rays are finely rough and tips end either bluntly sharp or rounded. Pentactins often have a short stub developed as the sixth ray. Atrialia are regular hexactins (70%), pentactins (30%), and a few stauractins with the same ornamentation and ray tips as dermalia ( Fig. 22B View FIGURE 22 ). Diactins ( Fig. 22C View FIGURE 22 ) are lightly curved, usually without swelling at the axial cross, and smooth except for roughened tips ending as the previous megascleres; they are relatively thin, maximum about 20 µm. Of the two diactine groups examined ( Table 11) overlap is great but parenchymal and atrial diactins are generally longer than hypodermal diactins.
(dimensions in µm unless otherwise indicated).
Microscleres consist of discoctasters, oxyhexactins, and hexasters; discohexasters are absent. Discoctasters ( Fig. 22D View FIGURE 22 ) are moderate size with basal half of the primary rays smooth and only the distal half finely rough. Each of the eight primary rays carries 4–10 slightly spreading, straight or slightly curved, rough terminal rays ending in marginally serrate discs ( Fig. 22E View FIGURE 22 ). The six interradial knobs on the original cube faces are well developed in octasters from both surfaces ( Fig. 22F View FIGURE 22 ). Discoctasters from the dermal side are much smaller than those from the atrial plus parenchymal areas ( Table 11) and probably represent distinct size classes, but there is considerable overlap and there are no other differences between them (primary to secondary ray lengths are 1.02 for dermal and 0.98 for atrial plus parenchymal). Oxy-tipped microscleres are thin and fragile ( Figs. 22G–I View FIGURE 22 ) with short primary rays sometimes subsumed by a swollen centrum, but locations of primary rays always remain clear. They are absent from the dermal surface tissues but abundant in atrial and parenchymal regions. Their forms are mainly hemioxyhexasters (80%), oxyhexasters (10%), and oxyhexactins (10%). Most, but not all, bear reclined spines on their terminal rays that increase in length and density basally; these feather-like basal spines form a conspicuous (in SEM, not LM) dense fringe around bases of terminal rays of hexasters or rays of hexactins ( Figs. 22J, K View FIGURE 22 ), while primary rays and exposed centrum surfaces are smooth and without ornaments. In addition, proclined barbs of various lengths occur commonly around the bases of hexaster terminal rays ( Fig. 22J View FIGURE 22 , arrowheads); they are interpreted to be short undeveloped terminal rays. Hexasters bear 1–4 terminals per primary ray and, as is common, hexasters are smaller than hexactins.
Etymology. The species name is formed in honour of Vladimir Koltun, who produced the basic reference work to North Pacific Hexactinellida, besides his extensive other publications on sponges.
Remarks. There are presently only six species/subspecies in Acanthascus sensu stricto. Although the new specimen differs from all known species/subspecies by several characters, it is expedient to list the most obvious. It is excluded from A. cactus Schulze, 1886 and A. pachyderma Okada, 1932 (both with tetractine dermalia) by its possession of mainly pentactine dermalia. The greater length of dermalia tangential ray length in the new form (130–178–209 µm) excludes it from A. alani alani Ijima, 1898 (125–175 µm), A. mitis Koltun, 1967 (110–140 µm), A. platei Schulze, 1899 (76–132–172) and A. sacculus Hernandez, 1932 (105–161 µm); the mean of the new form exceeds the greatest range in all four of those noted. Qualitative differences include presence of microdiscohexasters in A. alani alani , A. platei and A. sacculus (absent in the new form) and presence of distinctive hemioxyhexasters with one elongate axis bearing 4–5 terminal rays in one plane in A. mitis (no such spicules in the new form). This leaves only A. alani profundum Koltun, 1967 as a potential assignment for it. The Aleutian form, however, differs from A. a. profundum in oxy-microsclere diameter (98–199 vs 82–148 µm) and absence of the larger discoctaster class (diameter 330–425 µm) present in the latter. We conclude from these data that the Aleutian form is a new species, here designated Acanthascus koltuni , n. sp.
The new specimen has prompted detailed review of original descriptions of the two subspecies of A. alani . From this process, we propose raising A. alani profundum Koltun to species level, resulting in three closely related North Pacific species of Acanthascus with the following characters (among others). A. alani (subspecies discontinued) occurs shallower than 300 m, has microdiscohexasters, no spurs on oxyhexasters, no oxyhexactine microscleres, and discoctasters of 120–220 µm diameter. A. profundum Koltun (n. comb. for A. alani profundum ) occurs in deeper water (> 2000m), has no microdiscohexasters, has spurs on oxyhexasters, has oxyhexactine microscleres, and has two sizes of discohexasters, the larger class 330–425 µm in diameter. A. koltuni n. sp., also occurs in deep water (> 2000 m), lacks microdiscohexasters, has spurs on oxyhexasters, has oxyhexactine microscleres, and has two broadly overlapping discoctasters, but the larger is 171–286 µm in diameter. All three of these species share conspicuous spines at the base of oxyhexaster terminal rays, most likely derived from their common ancestor.
The only collected specimen was attached to a large boulder. Review of all video footage collected with the ROV 'Jason II' indicates that this is a rare species occurring singly on bedrock, mudstone, and large boulders at depths between 1446 and 2245 m.
Genus Staurocalyptus Ijima, 1897 View in CoL
Synonymy. Staurocalyptus Ijima, 1897: 53 View in CoL . Part of Rhabdocalyptus View in CoL - R. dowlingii Lambe, 1894: 37 ; 1900: 165; Schulze, 1897: 35 and R. roeperi Schulze, 1886: 51 View in CoL ; 1887 a: 158, 1897: 34. Part of Acanthascus Tabachnick 2002: 1447 View in CoL . Subgenus Staurocalyptus Tabachnick 2002: 1451 View in CoL .
Type species: Rhabdocalyptus dowlingi Lambe, 1894
Genus diagnosis. Acanthascinae with dermal spicules supported by a continuous hypodermal lattice formed of large pentactins, all of which lack thorns on their tangential rays (modification of Tabachnick 2002: 1451 for subgenus Staurocalyptus ).
Remarks. The group Staurocalyptus is restored to genus status as proposed above under Acanthascinae . Koltun (1967) designated R. dowlingi Lambe as type species of Staurocalyptus . Tabachnick's (2002) attempt to replace the type species with S. glaber is without merit. His 1999 application to ICZN (case 3104) produced no comment, opinion or declaration and thus his declaration of S. glaber (Tabachnick, 2002) contravened ICZN rules. Lambe's 1894 description was clear and his specimen remains available for inspection at the Canadian Museum of Nature. A new diagnosis is formed here, requiring presence of a supporting lattice of hypodermal pentactins, not simply the presence of a few scattered hypodermal pentactins, for inclusion in Staurocalyptus .

| RV |
Collection of Leptospira Strains |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Acanthascus koltuni
| Reiswig, Henry M. & Stone, Robert P. 2013 |
Staurocalyptus
| Ijima 1897 |
Staurocalyptus
| Ijima 1897: 53 |
R. dowlingii
| Lambe 1894: 37 |
R. roeperi
| Schulze 1886: 51 |
Staurocalyptus
| Tabachnick 2002: 1451: 1451 |
Rhabdocalyptus
| Tabachnick 2002: 1449 |
Acanthascus
| Tabachnick 2002: 1447: 1447 |