
Acanthostega (Coates, 1996)
|
publication ID |
https://doi.org/10.1093/zoolinnean/zlaa182 |
|
persistent identifier |
https://treatment.plazi.org/id/03D287C9-9D47-B076-E381-FA1E25EDD30F |
|
treatment provided by |
Plazi |
|
scientific name |
Acanthostega |
| status |
|
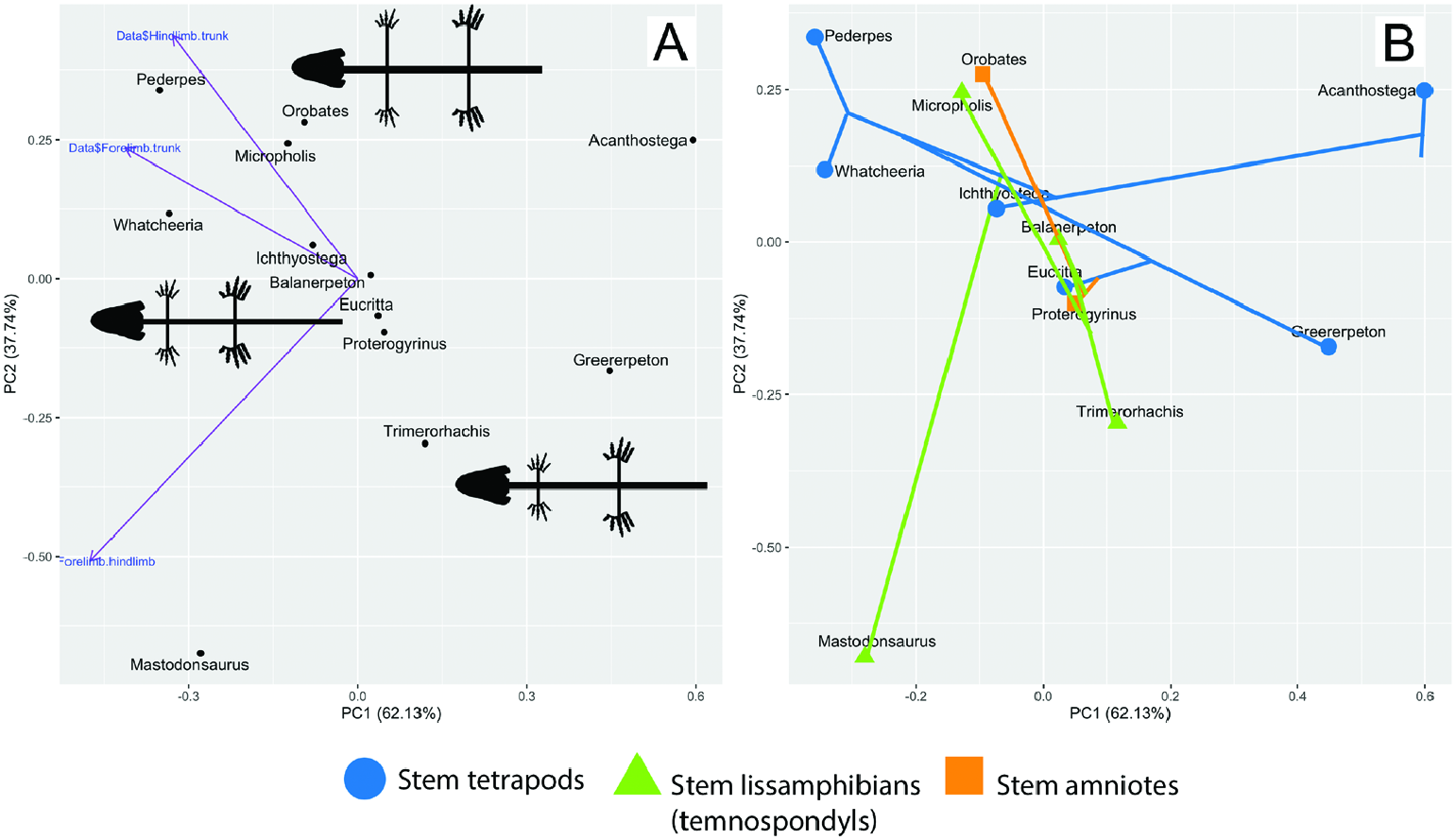
• Acanthostega / Trimerorhachis / Greererpeton (‘small’). • Balanerpeton / Proterogyrinus / Eucritta (‘medium’). • Whatcheeria / Pederpes / Micropholis / Orobates (‘large’).
Importantly, from these groups (albeit from a small sample size) there is no clear correlation between proportional limb size and inferences of terrestriality.
These numerical results are visualized in the most taxonomically inclusive permutation of the PC analysis ( Fig. 32 View Figure 32 ). Due to difficulty in obtaining reliable limb measurements for Seymouria , it was excluded from this permutation, though the measurements are reported in Table 2. Three morphotypes are discernible: those with large, equal-length limbs ( Micropholis , Orobates, Pederpes , Whatcheeria ); those with medium-length, medium-disparity limbs ( Balanerpeton , Eucritta , Ichthyostega , Proterogyrinus ); and those with small, medium-to-high-disparity limbs ( Acanthostega , Greererpeton , Trimerorhachis ). There is no discernible phylogenetic clustering. All three morphotypes are represented by both stem tetrapods and crown tetrapods. Notably, Whatcheeria and Pederpes are the stem tetrapods that converge most closely with the terrestrially adapted crown tetrapods in terms of limb proportions. Although these results signal that multiple tetrapod lineages converged on similar limb proportions, extrapolations to convergent function are less clear.
FUNCTIONAL ANATOMY
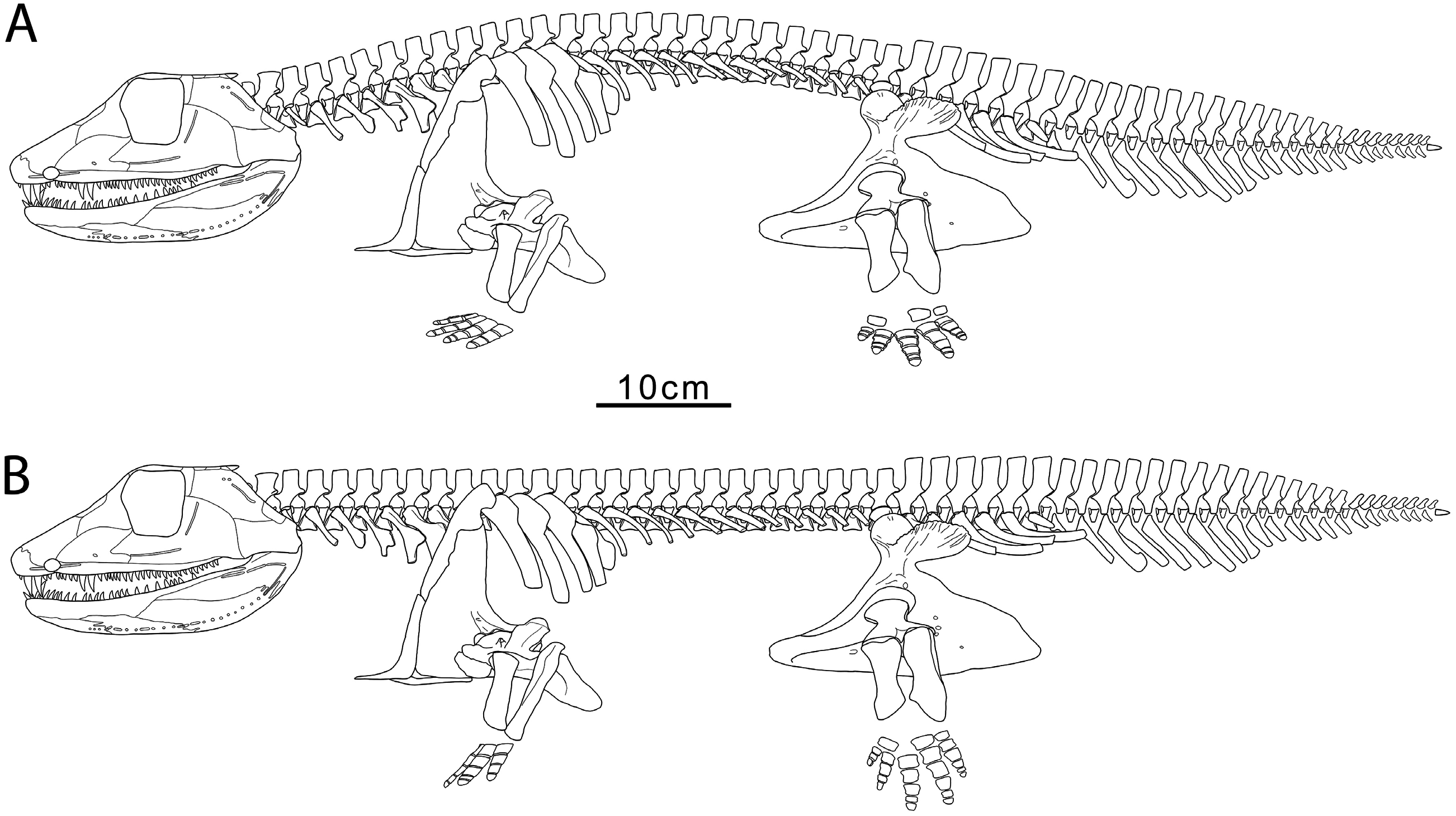
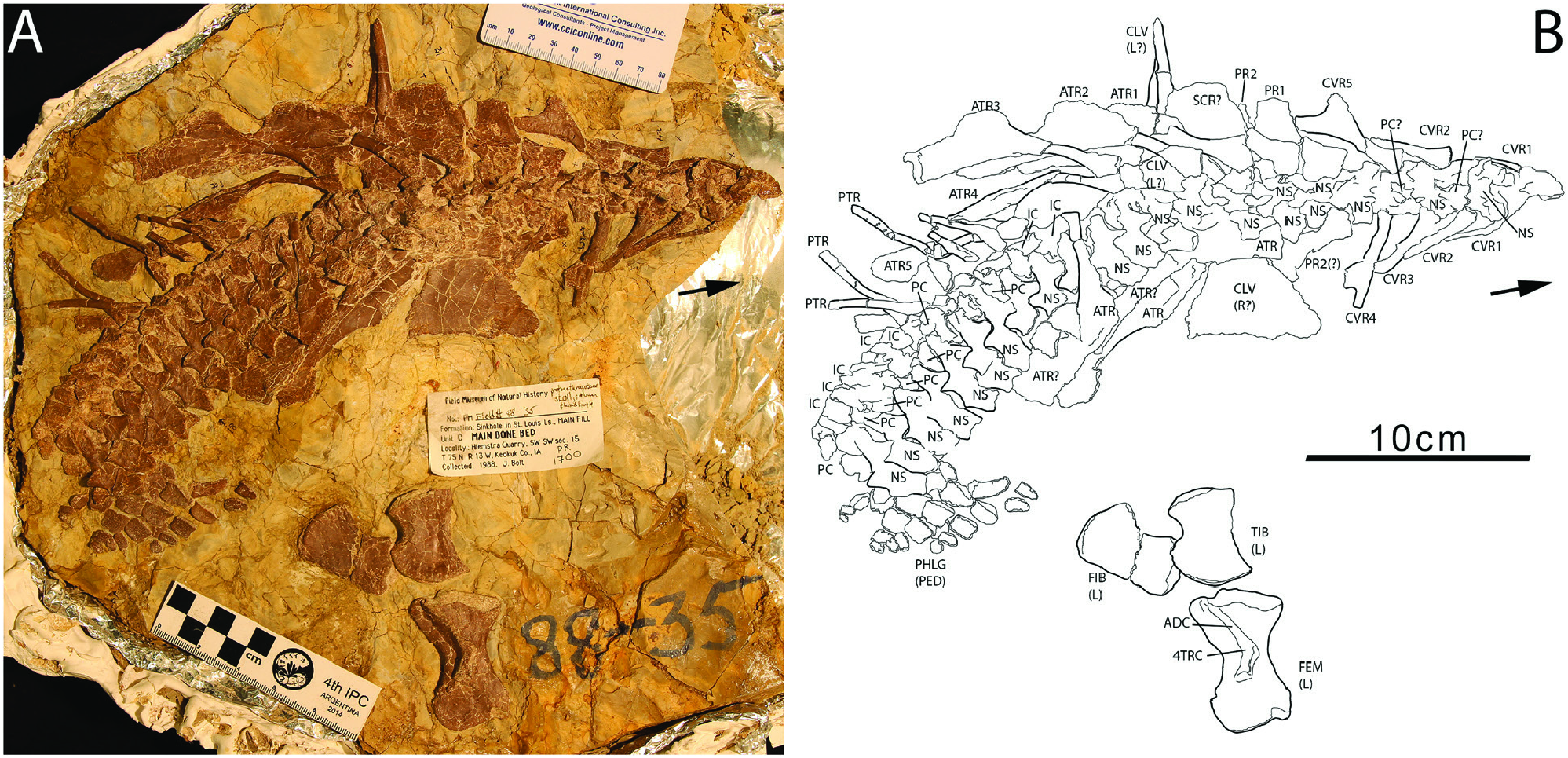
The Whatcheeria postcranium is more of a unique collection of plesiomorphies than a suite of novel characteristics. The uncinate processes of the ribs are particularly large, but uncinate processes are widely distributed across early tetrapods ( Coates, 1996; Clack & Finney, 2005). The humerus, with plesiomorphic L-shape and set of processes, is distinguished by size and proportions – including a massive entepicondyle – rather than its derived organization. The wrist and ankle are barely ossified, in common with most other early tetrapods. Indeed, the ankle is less ossified than in Acanthostega or Greererpeton , which were firmly aquatic and are traditionally recovered as stemward and crownward of Whatcheeria , respectively. Furthermore, the phalanges are short and broad, resembling those of the thoroughly ossified pes in the hindlimb paddle of Ichthyostega . In summary, rather than ongoing innovation, these morphologies seem more consistent with the phenomenon of constrained character space or character exhaustion ( Ruta et al., 2006; Wagner et al., 2006; Bernardi et al., 2016), earlier than might be expected in the initial evolutionary radiation of limbed tetrapods.
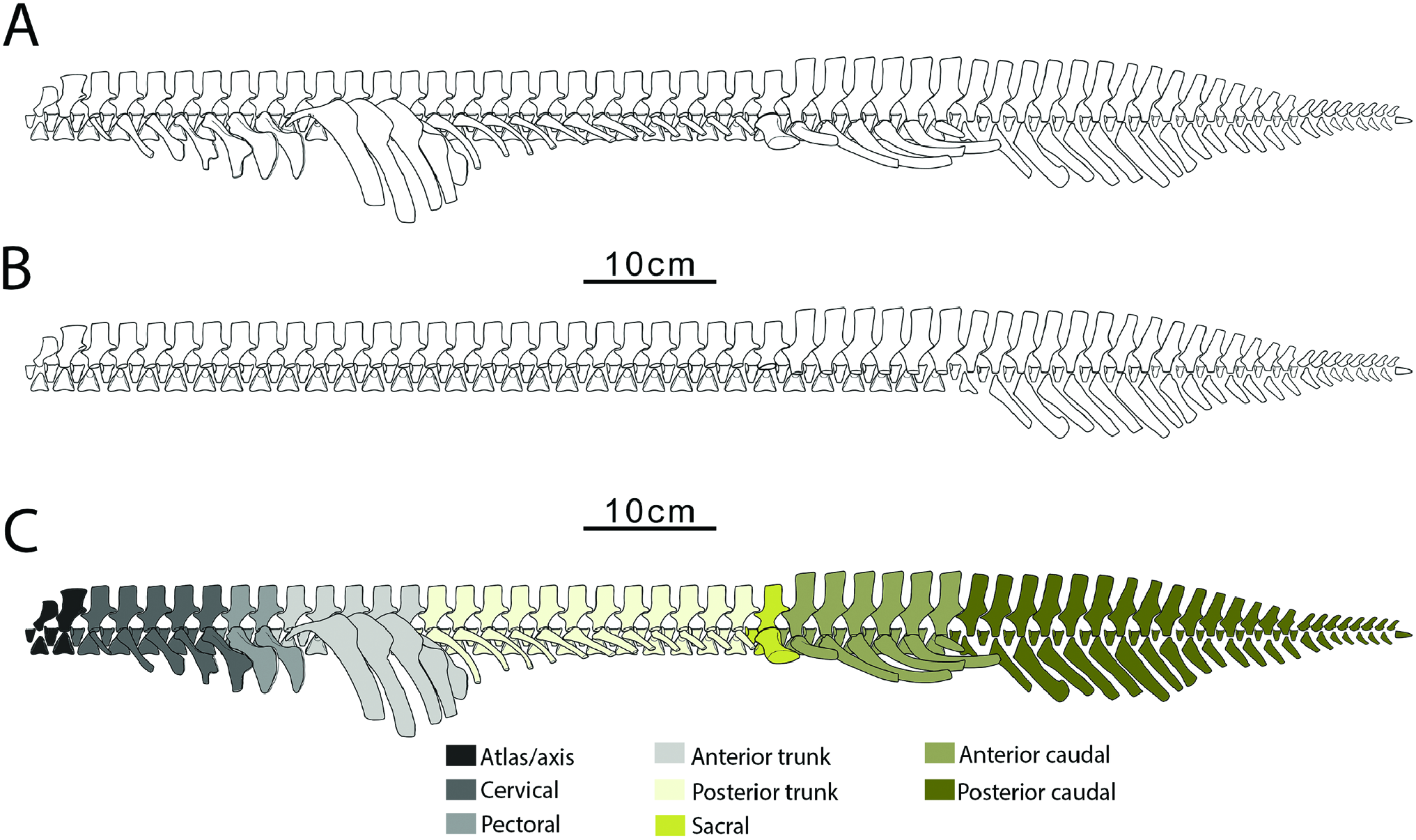
Whatcheeria , like Eryops and Ichthyostega , has a strongly regionalized ribcage with large uncinate processes on the elongate anterior trunk ribs. In all three taxa this arrangement was likely associated with substantial shoulder musculature. The shorter posterior trunk ribs of Whatcheeria are most likely plesiomorphic (cf. Acanthostega, Coates 1996 ). Notably, these ribs are not reduced as in the East Kirkton ‘anthracosauroid’ Eldeceeon Smithson, 1994, which has long, anterior trunk ribs but lacks posterior trunk rubs altogether ( Smithson, 1993; Ruta et al., 2020).
Increased shoulder musculature accords well with the extensive forelimb musculature implied by the morphology of the humerus and ulna. The limb would have been most powerful in retraction, with protraction being the recovery stroke. The shape of the humeral head and glenoid would have limited pronation and supination. Similarly, the olecranon process and ectepicondyle might have reduced the ability of the forelimb to extend at the elbow ( Holmes, 1984). The forelimb would have provided stable, pectorallevel support, but with limited forelimb extension or rotation, walking would have required retraction to lift the forelimb and lateral flexion of the axial column to swing it forwards. This was likely assisted by the short posterior trunk ribs. Kinematic study of terrestrial locomotion in extant Pleurodeles Michahelles, 1830 View in CoL ( Karakasiliotis et al., 2013) found large amounts of lateral flexion occurred in the posterior trunk and the distal half of the tail. Similarly, robotic biomechanical simulations of Orobates ( Nyakatura et al., 2019) , based on trackways and skeletal evidence, estimate that greatest lateral bending occurred approximately halfway along the trunk, with additional lateral bending at the base and midpoint of the tail. Whatcheeria lacks the morphological adaptations to the posterior trunk (‘lumbar’) vertebrae hypothesized to limit lateral (and dorsoventral) flexion in Ichthyostega ( Ahlberg et al., 2005) . Therefore, despite their similarities, Ichthyostega likely had a different locomotor strategy than Whatcheeria , possibly one unique among early tetrapods ( Ahlberg et al., 2005; Pierce et al., 2012).
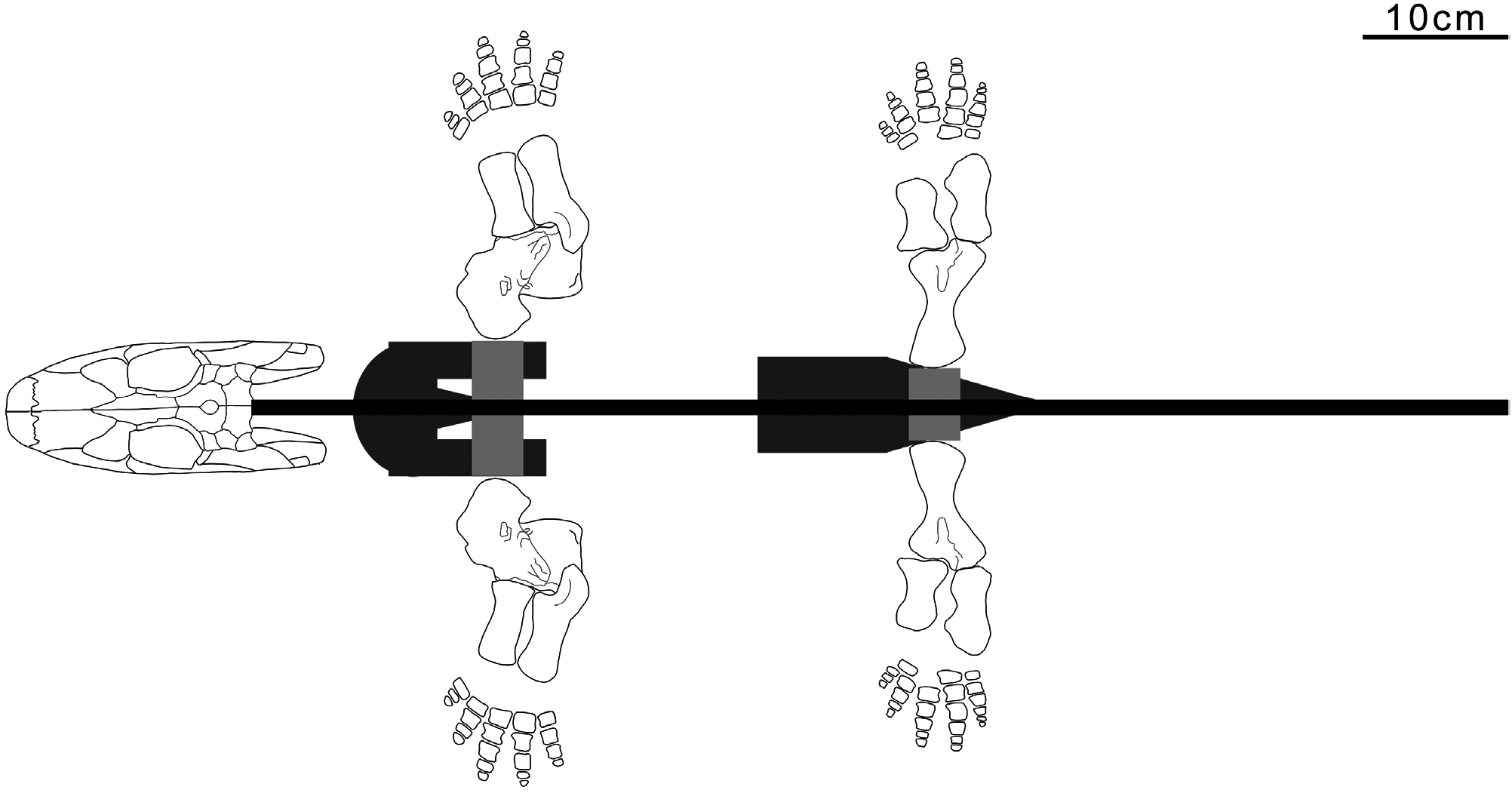
The squat phalanges of Whatcheeria are dissimilar to those of (putatively) terrestrial taxa, which tend to be waisted with ventral grooves to accommodate ligaments to assist grasping ( Clack & Finney, 2005). However, like Pederpes , the asymmetric pedal phalanges imply an anteriorly oriented foot ( Figs 4 View Figure 4 , 25 View Figure 25 , 31 View Figure 31 ), which in turn is a morphology associated with a walking gait (whether underwater or on land) ( Clack & Finney, 2005). The significance of the breadth of the phalanges is unclear. These might have enhanced support (in or out of the water), or served as paddle-skeleton, acknowledging that broad phalanges occur in the paddles of extinct and extant tetrapods.
Limb length relative to body length ( Figs 2 View Figure 2 , 31 View Figure 31 , 32 View Figure 32 ; Table 2) and limb robustness in Whatcheeria suggest an increased emphasis on appendicular locomotion and support compared to other early tetrapods ( Fig. 32 View Figure 32 ; Table 2). The tail lacks the radials and large neural and hemal spines of aquatic early tetrapods, suggesting it was less important for locomotion (although the extent of any soft-tissue fin is completely unknown).
As in Proterogyrinus ( Holmes, 1984) , it is unlikely that Whatcheria could pull its limbs flush with the body; they would have protruded during swimming. Absence of an internal trochanter and presence of a broad fourth trochanter and well-developed femoral adductor crest suggest an emphasis on the muscles elevating the body, and de-emphasis on those associated with moving the limb in a paddle-like action ( Panchen & Smithson, 1990). The morphology of the pelvic symphysis indicates that the pelvic girdle was strong and well buttressed. Extant analogues for swimming in Whatcheeria are difficult to identify, although the duckbilled platypus, Ornithorhynchus anatinus (Shaw, 1799) , provides an intriguing model. The platypus uses a rowing stroke, moving the forelimbs either in conjunction or alternately ( Howell, 1937; Fish et al., 1997; Fish, 2000). Like Whatcheeria , platypuses have a large and well-developed olecranon process on the ulna (with the caveats that the platypus also uses its forelimbs for digging, and that the elbow joint of Whatcheeria was not very mobile). Biomechanical research on swimming mammals shows that drag-based swimmers, such as platypuses, generate greater thrust at low speeds, increasing manoeuvrability ( Fish et al., 1997; Fish, 2000; Walker & Westneat, 2000). Such capability would be useful in the context of aquatic predation. It might also have contributed to functional trade-offs needed to maintain swimming capability in the context of largely aquatic biotas, such as that of Delta ( Bolt et al., 1988; Snyder, 2006; Bolt & Lombard, 2010), even as appendicular skeletal morphologies became increasingly adapted for occasional terrestrial support and locomotion.
The elongate neck of Whatcheeria is the most striking feature revealed by the present reconstruction ( Figs 2 View Figure 2 , 3 View Figure 3 ). Neck length must have increased head mobility, relative to the primitive condition wherein the pectoral girdle is close behind the cheek (e.g. Ichthyostega and Pederpes ). Such mobility would have reduced the need to move the entire body to track, grasp and manipulate prey. The increased space between the skull and pectoral girdle would also have allowed more room for jaw depression, increasing gape and throat volume. However, estimates of capacity are difficult without a preserved ceratohyal.
The unusual combination of morphologies present in Whatcheeria complicates the inference of its life habits. Whatcheeria appears to have been capable of walking, and perhaps was able to undertake limited terrestrial excursions. However, it is unlikely that Whatcheeria spent most or much of its time on land, given the presence of the cranial sensory canals and lack of robust wrist and ankle joints. Moreover, the Delta fauna is dominated by aquatic taxa, so Whatcheeria likely fed mostly or solely in the water. An aquatic walking gait, as hypothesized for Seymouria ( White, 1939) and Proterogyrinus ( Holmes, 1984) , seems plausible in Whatcheeria . Indeed, Whatcheeria may be a tetrapod specialized for walking in shallows, perhaps adapted for negotiating vegetation-choked, marginal habitats. This kind of locomotion was likely important and persistent within a range of water–land transition(s).
RE- EVALUATION OF WHATCHEERIID TAXA AND SPECIMENS

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Acanthostega
| Otoo, Benjamin K A, Bolt, John R, Lombard, R Eric, Angielczyk, Kenneth D & Coates, Michael I 2021 |
Acanthostega
| , Coates 1996 |
Pleurodeles
| Michahelles 1830 |