
Pseudolycoriella jejuna (Edwards, 1927)
|
publication ID |
https://doi.org/10.11646/zootaxa.4707.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:3C00FB35-708D-4FF0-94C2-B15BD2A1F37A |
|
persistent identifier |
https://treatment.plazi.org/id/03D43F59-3676-FFE9-FF32-FC34346E3A84 |
|
treatment provided by |
Plazi |
|
scientific name |
Pseudolycoriella jejuna |
| status |
|
Pseudolycoriella jejuna View in CoL complex
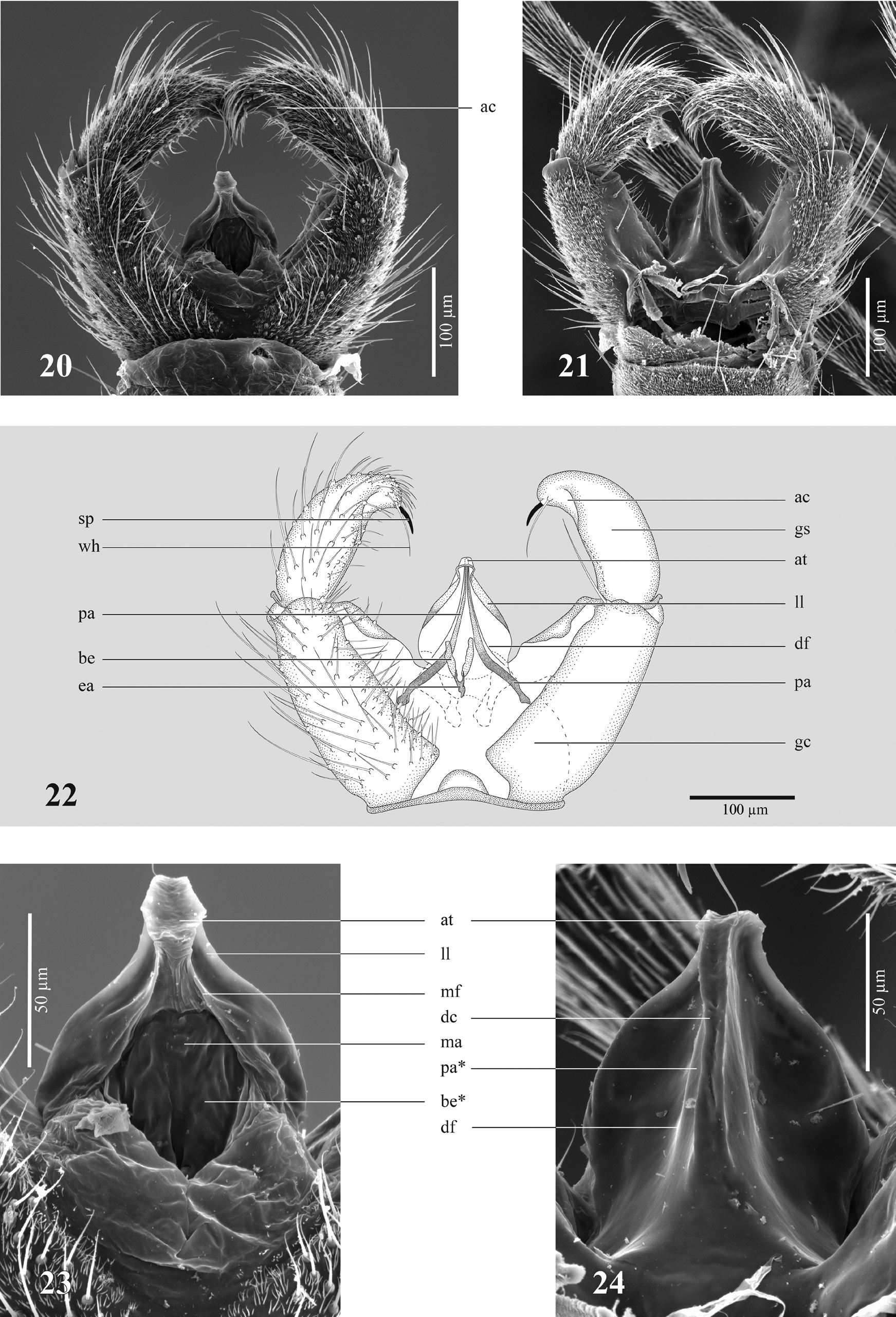
The Psl. jejuna complex is a group of thirteen closely related and cryptic species. These species share a complex and synapomorph structure of their hypopygia. In Figures 20–22 a View FIGURES 20–24 hypopygium of Psl. tewaipounamu is depicted. The gonostylus ( gs) is elongated and curved with an inwardly bent apex. The apex possesses one spine ( sp) and a whiplash hair ( wh) and—in most species—on the inward part a cavity ( ac). The tegmen is highly derivative compared with the tegmen of typical sciarids. Its general shape is onion-shaped: more or less roundish with an apical contraction ( at). Adjacent to the apical contraction the tegmen has two lateral lobes ( ll) on the ventral side, which can easily be observed in light microscopy. In the middle of the ventral side of the tegmen a membranous area ( ma) bordered by two median folds ( mf) can easily be observed by using scanning electron microscopy. In light microscopy these delicate structures are hardly visible. At this membranous area a bulge of the base of the ejaculatory apodeme ( be) can be observed. An area of teeth is absent. In light microscopy, the parameral apodemes ( pa) do not fuse medially. Instead they are stretched to the apex of the tegmen and form the apical contraction ( at). On the dorsal side of the tegmen the parameral apodemes protrude and form a dorsal carina ( dc). This carina is bent around the apex and ends subapically on the ventral side of the tegmen. The bulges of the parameral apodemes are accompanied by a dorsal fold ( df), which can also be observed in light microscopy. The phylogeny of this complex is shown in Figure 61 View FIGURE 61 .
Discussion. The species of the Psl. jejuna complex can be assigned to the Psl. bruckii group. The monophyly of this complex is strongly supported by genetics and can also be demonstrated by some autapomorphic morphological characters such as the extraordinary tegmen, the reduction of the number of gonostylar spines, and the secondary absence of an area of teeth. Due to this unique character set, all species of the Psl. jejuna complex can easily be recognized. Only Psl. fuscorubroides (Mohrig & Blasco-Zumeta) from Spain has a similar tegmen (Mohrig & Blasco- Zumeta 1996). It also possesses a dorsal carina that derives from the parameral apodemes. However, there are differences: the parameral apodemes are fused and not separated by a gap, as in the Psl. jejuna complex, and the carina in Psl. fuscorubroides arises vertically in the middle base of the tegmen. Furthermore, Psl. fuscorubroides possesses a large area of conspicous teeth ( Mohrig & Blasco-Zumeta 1996). Several other structures of the hypopygium are also disimilar. For instance, the gonostylus of Psl. fuscorubroides is armed with two short spines and has a conspicuous lobe on the dorsal margin and therefore differs significantly from the gonostyli of the Psl. jejuna complex. It is not clear whether the dorsal carina of the tegmen is a convergent or a homologous character. Maybe Psl. fuscorubroides is closely related to the Psl. jejuna complex, but it certainly does not belong to the crown group of the Psl. jejuna complex, since the absence of an area of teeth can be regarded as a synapomorpy of the New Zealand Psl. jejuna complex members. In other words, the hypothesis of a monophyly of the New Zealand representatives of the Psl. jejuna complex cannot be rejected.
The occurrence of two atavisms among the specimens of the Psl. jejuna complex confirms the above character polarisation regarding the single gonostylar tooth and the absence of the area of teeth. One paratype of Psl. dagae has two spines on the left gonostylus while the right one only bears one. This atavistic state is unique among all 233 investigated specimens belonging to the Psl. jejuna complex and indicates that the species of the Psl. jejuna complex derivate from an ancestor that bears two spines at the gonostylus, like the extant species of the Psl. bispina complex or the Psl. zealandica complex. The second detected atavism occured in two paratypes of Psl. whena , where an area of the teeth is present, although to a varying degree—one paratype shows only five tiny teeth, the other twelve.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |