
Aonidella cf. dayi Maciolek
|
publication ID |
https://doi.org/10.11646/zootaxa.3786.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:3388BBBF-B3C6-4F34-9E76-E287CD335933 |
|
DOI |
https://doi.org/10.5281/zenodo.5630106 |
|
persistent identifier |
https://treatment.plazi.org/id/03D4B72B-6160-FF98-F7D1-A676FE3F4555 |
|
treatment provided by |
Plazi |
|
scientific name |
Aonidella cf. dayi Maciolek |
| status |
|
Aonidella cf. dayi Maciolek View in CoL in López-Jamar, 1989
( Figures 2 View FIGURE 2 –5)
Prionospio (Minuspio) cf. cirrobranchiata Day, 1961 View in CoL : Johnson, 1984: 6 –59, figures 6–47, 6–48. Prionospio cirrobranchiata Day, 1973: 73 View in CoL , figure 10g –k. (not the description but material examined) Aonidella dayi López-Jamar, 1989: 107 View in CoL –110, figures 1–2.—? Imajima, 1992: 57 –60, figures 1–2.— Maciolek, 2000: 531 –533,
figure 2.
Type material: Aonidella dayi Maciolek in López-Jamar, 1989: Paratypes: NW Atlantic Ocean, North Carolina, off Beaufort, 34°19'N, 75°56 'W, 130 m, coll. J. H. Day, 5 paratypes (all af) ( USNM 43154, as P. cirrobranchiata ).
Non– type material. Aonidella dayi Maciolek in López-Jamar, 1989: SW Atlantic Ocean: Brasilien, St. Y 1, van Veen, 60 m, 11 Jun 2002, 2 af ( SMF 14000), stn. 6677, van Veen, 12 Jan 1998, 2 af ( SMF 13993). NE Atlantic Ocean, Canary Islands: 27°13'N, 15°41 W, Discovery sta. 6711, epibenthic sled, 3301 m, 20 Mar 1968, coll. H. L. Sanders, 2 af ( USNM 80479).— Aonidella cf. dayi Maciolek in López-Jamar, 1989: Mid-Atlantic Expedition 2003/2004, cruise METEOR ME 60-1, Seine Seamount: 33° 49.1'N, 14° 21.9'W, station 754, multicorer, depth 209 m, 04-Dec-2003, formalin, 4 complete, 1 af ( SMF 16714); 33° 48'N, 14° 22'W, station 755, multicorer, depth 207 m, 04-Dec-2003, formalin, 1 af ( SMF 16718), 2 af ( SMF 16719); 33° 46.1'N, 14° 22'W, station 756, multicorer, depth 178 m, 04-Dec-2003, formalin, 2 complete, 1 af ( SMF 16710), 1 complete ( SMF 16711), 1 complete ( SMF 16712), 1 complete ( SMF 16713), 1 complete, 2 af ( SMF 16715), 1 complete ( SMF 16716), 1 complete ( SMF 16717), 4 complete, 1 af ( SMF 16720). Expedition DIVA 3, cruise METEOR ME 79-1: Irving Seamount: 31°56.98'N, 027°56.63'W, station 682, grab, depth 281.2 m, 21-Aug-2009, formalin: 1 complete ( SMF 23376), 2 complete, 1 af (ZSRO-P2339); 31°56.98'N, 027°56.66'W, station 683, grab, depth 281.1 m, 21-Aug- 2009, formalin: 1 complete ( SMF 23373), 96% ethanol: 6 af (ZSRO-P2364), 1 af ( SMF 23402); 31°56.98'N, 027°56.67'W, station 684, grab, depth 280.8 m, 21-Aug-2009, formalin: 5 af (ZSRO-P2354), 4 af ( SMF 23390), 1 af SEM ( SMF 23391); 31°56.98'N, 027°56.71'W, station 687, grab, depth 280 m, 21-Aug-2009, formalin: 10 af (ZSRO-P2336), 4 af (ZSRO-P2337); 31°56.97'N, 027°58.33'W, station 688, trawl, depth 269.3 m, 21-Aug-2009, formalin: 2 af (ZSRO-P2353); 31°56.96'N, 027°56.51'W, station 679, grab, depth 282.5 m, 21-Aug-2009, 96% ethanol: 1 complete, 2 af ( SMF 23401), 2 af (ZSRO-P2372); 31°56.96'N, 027°56.55'W, station 680, grab, depth 281.9 m, 21-Aug-2009, formalin: 12 af (ZMH-P27568), 1 complete ( SMF 23377); 31°56.94'N, 027°55.90'W, station 678, grab, depth 283.2 m, 21-Aug-2009, formalin: 2 complete, 1 af (ZSRO-P2335), 2 complete, 4 af (ZMH- P27567), 3 af ( SMF 23371). Hyères Seamount: 31°26.40'N, 028°56.13'W, station 669, grab, depth 290.8 m, 20- Aug-2009, formalin: 2 af (ZSRO-P2355), 1 af (ZSRO-P2356), 96% ethanol: 2 af ( SMF 23403), 1 af (ZSRO- P2365); 31°26.40'N, 028°56.22'W, station 670, grab, depth 290.8 m, 20-Aug-2009, formalin: 1 af (ZMH-P27561); 31°26.40'N, 028°56.27'W, station 671, grab, depth 288 m, 20-Aug-2009, formalin: 1 complete ( SMF 23393), 4 af ( SMF 23392), 96% ethanol: (ZSRO-P2357); 31°26.40'N, 028°56.31'W, station 672, grab, depth 286.2 m, 20-Aug- 2009, formalin: 3 af (ZSRO-P2340), 1 complete, 2 af (ZSRO-P2344). Great Meteor Seamount: 30°00.04'N, 028°30.05'W, station 660, grab, depth 287.2 m, 19-Aug-2009, formalin: 4 af ( SMF 23369), 16 af (ZSRO-P2333), 2 complete, 14 af (ZMH-P27569), 96% ethanol: 2 af ( SMF 23406); 30°00.04'N, 028°30.05'W, station 661, grab, depth 287.4 m, 19-Aug-2009, formalin: 6 af ( SMF 23372), 1 af ( SMF 23383), 20 af (ZSRO-P2348), 96% ethanol: 8 af ( SMF 23399), 6 af (ZSRO-P2363), 1 af ( SMF 23400), 3 af (ZSRO-P2369); 30°00.04'N, 028°30.05'W, station 662, grab, depth 287.6 m, 19-Aug-2009, formalin: 1 complete, 6 af, 1 af SEM (ZSRO-P2329), 8 af (ZSRO-P2330), 96% ethanol: 6 af ( SMF 23408); 30°00.04'N, 028°30.05'W, station 663, grab, depth 287.1 m, 19-Aug-2009, formalin: 2 af (ZSRO-P2343), 4 af ( SMF 23381), 2 af (ZSRO-P2346), 96% ethanol: 1 complete, 2 af ( SMF 23398); 30°00.04'N, 028°30.05'W, station 664, grab, depth 287.9 m, 19-Aug-2009, formalin: 1 complete, 2 af (ZMH-P27563), 13 complete, 1 complete SEM, 31 af, 3 af SEM (ZMH-P27565), 2 complete, 4 af (ZSRO-P 2179); 30°00.00'N, 028°29.99'W, station 659, epibenthic sledge, depth 287.7 m, 19-Aug-2009, 96% ethanol: 3 af ( SMF 23409); 29°44.24'N, 028°24.54'W, station 657, epibenthic sledge, depth 285.5 m, 18-Aug-2009, 96% ethanol: 3 af (ZSRO-P2366), 1 af ( SMF 23410); 29°44.13'N, 028°24.91'W, station 656, epibenthic sledge, depth 283.3 m, 18- Aug-2009, 96% ethanol: 1 af ( SMF 23404), 3 complete, 2 af (ZSRO-P2367); 29°42.17'N, 028°22.96'W, station 650, grab, depth 289.2 m, 18-Aug-2009, formalin: 10 af ( SMF 23379), 1 af SEM ( SMF 23380), 96% ethanol: 2 af ( SMF 23407); 29°42.16'N, 028°22.99'W, station 649, grab, depth 289 m, 18-Aug-2009, formalin: 2 af ( SMF 23374), 1 af (ZSRO-P2338), 1 af ( SMF 23375); 29°42.09'N, 028°22.76'W, station 647, grab, depth 289.9 m, 18- Aug-2009, formalin: 1 complete, 3 af (ZSRO-P2352), 96% ethanol: 1 complete (ZSRO-P2358); 29°41.97'N, 028°22.62'W, station 646, grab, depth 292 m, 18-Aug-2009, formalin: 6 af (ZMH-P27566), 11 af (ZSRO-P2350), 96% ethanol: 1 af (ZSRO-P2371); 29°41.84'N, 028°22.55'W, station 645, grab, depth 295 m, 18-Aug-2009, 96% ethanol: 6 af (ZSRO-P2370); 29°41.72'N, 028°22.39'W, station 644, grab, depth 300.4 m, 18-Aug-2009, formalin: 2 af (ZSRO-P2332), 1 complete, 3 af (ZSRO-P2351); 29°41.59'N, 028°22.41'W, station 643, grab, depth 299.5 m, 18-Aug-2009, formalin: 1 af ( SMF 23384), 1 af ( SMF 23386); 29°41.51'N, 028°22.24'W, station 642, grab, depth 307.5 m, 18-Aug-2009, formalin: 1 af ( SMF 23382), 2 complete, 1 af ( SMF 23388), 96% ethanol: 1 af (ZSRO- P2362); 29°41.46'N, 028°22.14'W, station 641, grab, depth 313.3 m, 18-Aug-2009, formalin: 1 af ( SMF 23367), 3 af (ZSRO-P2345), 1 af SEM ( SMF 23389). Little Meteor Seamount: 29°38.19'N, 028°58.98'W, station 631, grab, depth 272.3 m, 17-Aug-2009, formalin: 2 af ( SMF 23365), 1 af SEM ( SMF 23366), 2 af ( SMF 23387); 29°38.19'N, 028°59.07'W, station 632, grab, depth 271.7 m, 17-Aug-2009, formalin: 3 af (ZSRO-P2347), 96% ethanol: 2 af ( SMF 23394); 29°38.18'N, 028°58.86'W, station 630, grab, depth 272.3 m, 17-Aug-2009, 96% ethanol: 2 af (ZSRO-P2359), 96% ethanol: 3 af ( SMF 23405); 29°38.18'N, 028°59.26'W, station 634, grab, depth 276.7 m, 17- Aug-2009, formalin: 1 af (ZMH-P27562), 2 af (ZMH-P27570); 29°38.16'N, 028°59.17'W, station 633, grab, depth 272.2 m, 17-Aug-2009, formalin: 1 af (ZSRO-P2341), 1 complete ( SMF 23378), 1 af ( SMF 23385); 29°36.32'N, 028°59.93'W, station 625, grab, depth 276 m, 17-Aug-2009, formalin: 2 af (ZMH-P27564), 3 af ( SMF 23368); 29°36.32'N, 029°00.06'W, station 626, grab, depth 277.7 m, 17-Aug-2009, formalin: 6 af (ZSRO-P2334), 2 af ( SMF 23370); 29°36.29'N, 028°59.34'W, station 622, grab, depth 267.5 m, 17-Aug-2009, 96% ethanol: 7 af ( SMF 23396), 2 af (ZSRO-P2361), 1 complete ( SMF 23397); 29°36.29'N, 028°59.68'W, station 624, grab, depth 274.2 m, 17-Aug-2009, 96% ethanol: 2 af ( SMF 23395), 96% ethanol: 18 af (ZSRO-P2360); 29°36.28'N, 028°59.53'W, station 623, grab, depth 270.9 m, 17-Aug-2009, formalin: 6 af ( SMF 23364); 29°36.27'N, 028°58.81'W, station 619, grab, depth 268.2 m, 17-Aug-2009, formalin: 1 af (ZSRO-P2331), 4 af (ZSRO-P2342), 1 af (ZSRO-P2349), 96% ethanol: 1 af (ZSRO-P2368).
Additional material examined: Aonidella cirrobranchiata ( Day, 1961) : NE Atlantic Ocean, Bay of Biscay, 46°17.5' N, 04°45.2’ W, R/V Sarsia, sta. 63, epibenthic sled, 24 Jul 1967, 1336 m, coll. H. L. Sanders, 1 af ( USNM 80482).— Aonidella insolita comb. nov. (Greaves, Meißner & Wilson, 2011): Paratypes: Indian Ocean, Shark Bay region, L6, Stn SS07/2005 81, 25º57.583′ S, 112º15.417′ E, 28 Jan 2005, 393 m, 2 af, 1 mf, (ZSRO-P2062 ex MV F 110727), 2 af (ZMH-P25871 ex MV F 110727); non-type material: Indian Ocean: Kalbarri region, L8 400, Stn SS07/2005 88, 27º55.833′ S, 113º4.683′ E, 29 Jan 2005, 416 m, 1 complete, 6 af (MV F 110606); Shark Bay region, L6, Stn SS07/2005 81, 25º57.583′ S, 112º15.417′ E, 28 Jan 2005, 393 m, 1 complete, 1 af (MV F 160686); Point Cloates region, L3, Stn SS07/2005 69, 22º51.533′ S, 113º19.617′ E, 27 Jan 2005, 451 m, 1 complete (MV F 110618); Carnarvon region, L5, Stn SS07/2005 76, 24º35.183′ S, 112º15.233′ E, 27 Jan 2005, 405 m, 1 complete, 1 af (MV F167436); Zyutdorp region, L7 400, Stn SS07/2005 85, 27º10.05′ S, 112º46.683′ E, 29 Jan 2005, 375 m, 5 af (MV F158685); Abrolhos region, T3 400, Stn SS07/2005 98, 28º59.433′ S, 113º45.967´E, 30 Jan 2005, 388 m, 1 complete, 1 af (MV F167416).
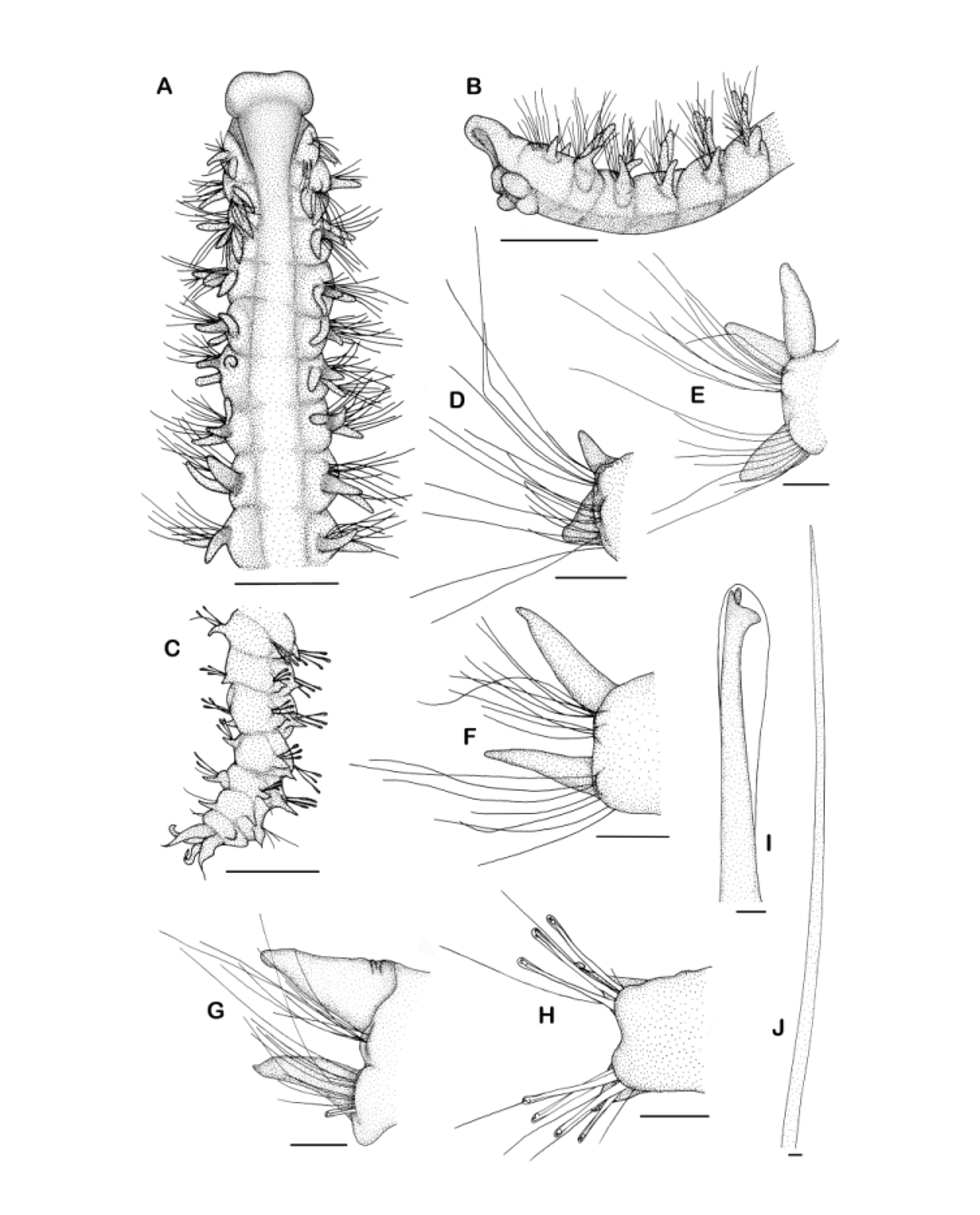
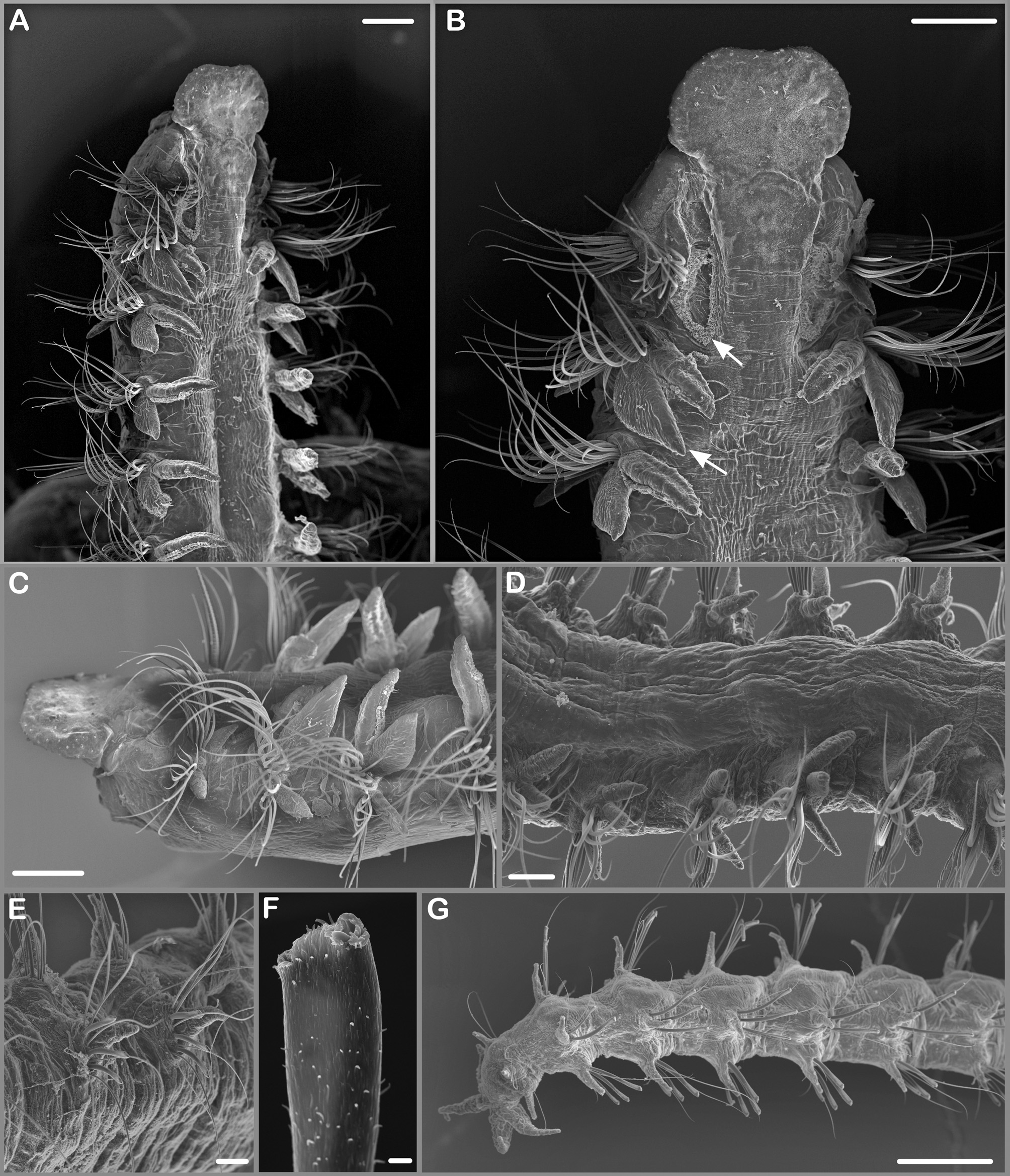
Diagnosis. Prostomium with broadly rounded, distinctly projecting forward over peristomium; posterior prostomial margin not discernable as completely fused with dorsum ( Figures 2 View FIGURE 2 A–B, 3A–C); occipital antenna absent. Nuchal organ as U-shaped ciliary band lateral to the fused prostomium and dorsum extending to the beginning of the second chaetiger ( Figure 3 View FIGURE 3 A–B). Metameric dorsal ciliated organs absent. Interparapodial lateral pouches absent. Dorsal branchiae from chaetiger 2 to maximally chaetiger 17 ( Figures 2 View FIGURE 2 A, 3A–D, 5 left). Notopodial postchaetal lamellae on chaetiger 2 enlarged in medium- and large-sized specimens ( Figure 3 View FIGURE 3 B). Hooded hooks with pair of small, somewhat perpendicular apical teeth above main fang present in noto- and neuropodia ( Figures 2 View FIGURE 2 C, I, H, 3F, 5 right). Rows of holes present on the dorsum of chaetigers of the branchiate region or adjacent to it (only observable with SEM, Figure 4 View FIGURE 4 ). Pygidium with 4 cirri of about equal length ( Figures 2 View FIGURE 2 C, 3G).
Description. Widest specimen 0.57 mm wide (at chaetiger 10), 17.5 mm long for 71 chaetigers. Prostomium with broadly rounded, slightly inflated anterior rim, distinctly projecting forward over peristomium, laterally separated from peristomium by furrow, posterior margin not discernable as completely fused with dorsum ( Figures 2 View FIGURE 2 A–B, 3A–C). Occipital antenna absent. Prostomium with two pairs of small red eyes arranged in trapezoid: anterior pair farther apart with eyes located just behind the inflated anterior rim, posterior pair close to the anterior pair, eyes close to the lateral prostomial margins; usually lost in formalin-fixed specimens but visible in ethanolfixed material. Peristomium separated from the prostomium, not well developed. Palps arising in front of the nuchal organs, reaching back to chaetiger 10 ( Figure 3 View FIGURE 3 B, palps lost in this picture but scar at former point of attachment discernable). Nuchal organ as U-shaped ciliary band lateral to the fused prostomium and dorsum reaching the beginning of chaetiger 2 ( Figure 3 View FIGURE 3 A–B). Metameric dorsal ciliated organs absent.
Dorsal branchiae present from chaetigers 2 to maximally chaetiger 17, last branchiate chaetiger dependent on specimen size, branchiae usually present to chaetiger 6 in small specimens, in very small specimens very few branchiae present ( Figures 2 View FIGURE 2 A, 3A–D, 5 left); branchiae easily lost. Branchiae cirriform, ciliated on outer margins, separate from notopodial lamellae throughout; first and last branchiae smallest, about half as long as notopodial lobe of the respective chaetiger; branchiae in between all of about the same length, in small specimens slightly shorter than notopodial lamellae, in medium-sized and large specimens as long as notopodial lamellae of the same chaetiger or longer, but even in largest specimens hardly touching above dorsum ( Figures 2 View FIGURE 2 A, E, 3A–D, 4A–C).
Parapodia on first and second chaetiger slightly more dorsally positioned than parapodia in following chaetigers ( Figure 3 View FIGURE 3 A). Both notopodial and neuropodial lamellae relatively small. On first chaetiger notopodial lamellae very small, short cirriform; neuropodial lamellae subtriangular with broad base, longer than notopodial lamellae ( Figure 2 View FIGURE 2 D). Second chaetiger with fully developed notopodial lamellae, often longest notopodial lamella of all, longer than branchiae of the respective chaetiger, leaf-like and prominent particularly in larger specimens ( Figures 3 View FIGURE 3 B, 4A–B); neuropodial lamellae subtriangular to cirriform, shorter than notopodial lamellae ( Figure 3 View FIGURE 3 C). From third chaetiger notopodial and neuropodial lamellae in branchiate chaetigers tapered to cirriform with notopodial lamellae sometimes slightly longer than neuropodial lamellae or both of equal length ( Figure 2 View FIGURE 2 E). Postbranchiate region with shorter lamellae with broader base in both rami, further decreasing in size towards the end of the body though in about 5–8 posteriormost chaetigers again slightly longer and cirriform ( Figures 2 View FIGURE 2 G–H, 3G). Praechaetal lamellae not developed but large specimens with little bulge stainable with methyl green in front of notopodial chaetae in some chaetigers of the branchiate region.
Interparapodial lateral pouches absent. Dorsal crests absent.
Two rows of holes present on the dorsum of some chaetigers of the branchiate region or adjacent to it (only observable with SEM, Figure 4 View FIGURE 4 ): starting on chaetigers 3–5 with few large holes (N=1–2) next to the branchial base, on succeeding chaetigers number of holes increasing to up to 10, then arranged in a straight row from the branchial base nearly all the way across the segment ( Figure 4 View FIGURE 4 A–C); from about chaetiger 6 second anterior group of holes present, again starting with few holes only, later with increasing number of holes (N max=7) describing an outward curved line from the notopodial base to the anterior intersegmental border ( Figure 4 View FIGURE 4 A–C); both rows of large holes often accompanied by irregular anterior and posterior rows of small holes ( Figure 4 View FIGURE 4 D). Additional single or very few large holes sometimes present laterally between parapodia of the branchiate region ( Figure 4 View FIGURE 4 A). Cilia originating from inside the large holes often present (impairing the observation of this character). First postbranchial chaetiger usually with very few large and some small holes next to the branchial base; on following chaetigers holes no longer present ( Figure 4 View FIGURE 4 E).
Chaetae in two rows, anterior and middle chaetigers with two rows of capillaries ( Figure 3 View FIGURE 3 C), until chaetigers 10–14 with granulations, afterwards smooth ( Figure 2 View FIGURE 2 J), until chaetigers 20–25 with narrow sheaths, thereafter without sheaths; anterior row of capillaries slightly shorter than posterior row. Anterior row of capillaries replaced by row of hooks ( Figure 3 View FIGURE 3 E) from chaetigers 15–22 in neuropodia and from chaetigers 19–37 in notopodia, first presence of hooks depending on specimen size (Figure 5 right); hooks hooded, tridentate, with pair of small, somewhat perpendicular apical teeth above main fang ( Figures 2 View FIGURE 2 I, 3F); 1–7 hooks in neuropodia and 1–6 hooks in notopodia ( Figure 2 View FIGURE 2 H). Posteriormost chaetigers with 3–4 long simple capillaries behind rows of hooks in neuro- and notopodia ( Figure 3 View FIGURE 3 G). Sabre chaetae usually absent but single large specimen from Seine Seamount with sabre chaeta in three posterior chaetigers (50th–52th) at the end of the fragment.
Pygidium with 4 cirri of about equal length, arranged as dorsal and ventral pair; cirri with thin elongated tips ( Figures 2 View FIGURE 2 C, 3G); anus terminal.
Pigmentation. Pigment not discernable in preserved specimens.
Methyl green staining pattern. Anterior inflated rim of the prostomium, peristomium, branchiae, and postchaetal lamellae most intensively stained. In larger specimens sometimes darker stained low praechaetal bulge in notopodia of branchiate chaetigers observable. Some specimens with metameric pairs of dark-blue dots on the ventrum from about chaetiger 8 to some mid-body chaetigers.
Ecology. Aonidella cf. dayi was found on seamounts of the NE Atlantic in coral and crushed shell gravel in about 178–300 m of depth.
Geographical distribution. NE Atlantic Ocean: Great Meteor Seamount, Little Meteor Seamount, Irving Seamount, Hyères Seamount, Seine Seamount. The species may also occur in the Mediterranean Sea, off the Atlantic coast of the Iberian Peninsula and in deep waters south of the Canary Islands (see Remarks).
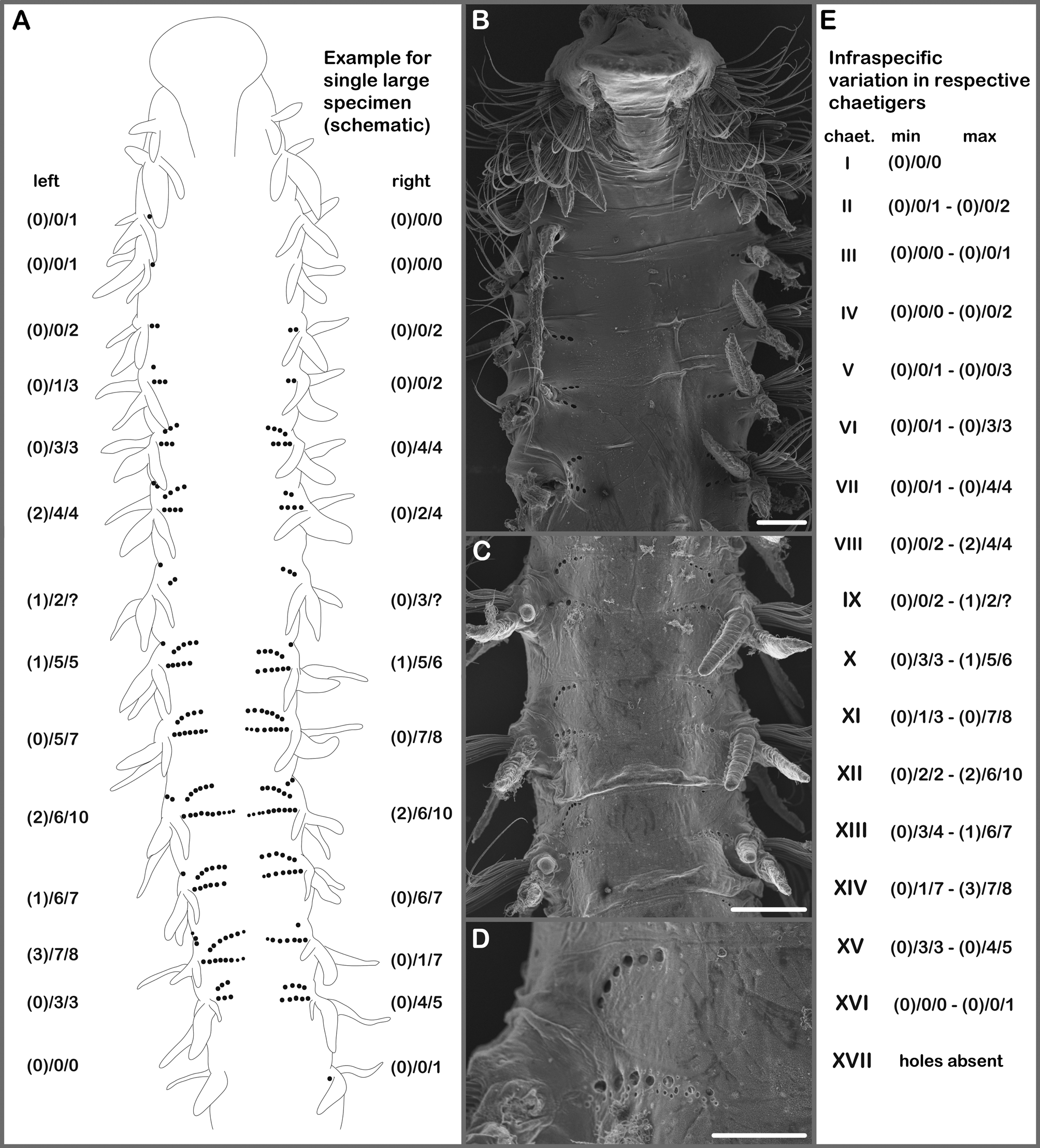
Remarks. Aonidella specimens found on the NE Atlantic seamounts are very close to Aonidella dayi Maciolek in López-Jamar, 1989. The amended description of A. dayi by Maciolek (2000) was based on the examination of specimens mainly from NW Atlantic but also on few specimens from the Mediterranean Sea and from deep waters South of the Canary Islands. Maciolek (2000) already noticed that her description reports the presence of 10–12 pairs of branchiae whereas other authors reported up to 16 pairs of branchiae, e.g Imajima (1992) for specimens from Japan and López-Jamar (1989) for specimens from the Gulf of Cadiz. In specimens from the Atlantic seamounts the number of branchiae was found to be size-dependent and largest specimens usually to have branchiae up to chaetigers 12–17 (i.e. 11–16 pairs of branchiae) (Figure 5 left). Direct comparison of specimens from both sides of the Atlantic Ocean revealed that largest specimens (> 0.5 mm wide at chaetiger 10) from off Brazil and from off the U.S. coast have fewer pairs of branchiae compared with specimens of the same size from the Eastern Atlantic seamounts (11–13 pairs versus 14–16 pairs) (Figure 5 left). Also many small seamount specimens ( 0.18–0.5 mm wide) with 10–15 pairs of branchiae were found. Differences concerning the start of neuropodial hooks between specimens from the Eastern and Western Atlantic were less distinct (Figure 5 right).
For A. dayi View in CoL from the Western Atlantic the presence of hooks with 2–4 apical teeth was reported ( Maciolek 2000). In their taxonomic catalogue of the Spionidae View in CoL of the Grand Caribbean Delgado-Blas & Salazar-Silva (2011) list a record of Aonidella cirrobranchiata ( Day, 1961) View in CoL with the note, that the presence of this species in the Grand Caribbean is doubtful because specimens of Johnson (1984) (Gulf of Mexico) and Ibarzabal (1986) ( Cuba) have a slight medial indentation on the prostomium, four eyes instead of none, and hooded hooks with 2–4 teeth. It seems obvious that those records should rather be assigned to A. dayi View in CoL than to A. cirrobranchiata View in CoL , but again the presence of hooks with more than 2 apical teeth is reported. For specimens of A. dayi View in CoL from the Eastern Atlantic only the presence of hooks with 2 apical teeth is known (this study and López-Jamar, 1989).
Specimens of A. dayi View in CoL from different sides of the Atlantic (records from the Caribbean, off the U.S. coast, and off Brazil and records from the Eastern Atlantic seamounts, the Canary Islands, the Western European coasts and the Mediterranean Sea) might belong to different species. The number of branchiae as well as the presence of hooks with more than 2 apical teeth could be distinguishing characters. However, the material available for the different geographical regions (except the seamount areas) was not sufficient to prove this idea. Either the limited number of specimens or their bad condition impeded the observation of important traditionally used characters and also the study of newly discovered morphological features as the arrangement of dorsal holes. Further morphological and genetic studies are required to resolve the taxonomy of A. dayi View in CoL . Such studies should also include specimens from Japan (see Imajima 1992).
Aonidella dayi View in CoL is most similar to A. insolita View in CoL comb. nov. from off Western Australia. In a search for distinguishing characters we also examined specimens of A. insolita View in CoL comb. nov. including some paratypes and can add the following information to the original description: nuchal organs are U-shaped ciliary bands lateral to the fused prostomium and dorsum reaching the beginning of chaetiger 2, small praechaetal notopodial lamellae present from chaetigers 2–18 in larger specimens (> 0.4 mm wide at chaetiger 10) and slight praechaetal bulges stainable with methyl green in smaller specimens, anterior row of capillaries with granulations in chaetigers 1–14, capillaries limbate up to about chaetigers 21–26, afterwards simple, in neuropodia up to 5 and in notopodia up to 4 hooks replacing anterior row of capillaries, pygidial cirri of about equal length in juvenile and adult specimens (observation of unequal length of pygidial cirri in juvenile specimens by Greaves et al. (2011) might be explained if the observation was of an regenerated specimen). According to our findings there are not many differences between A. insolita View in CoL comb. nov. and A. dayi View in CoL . The most obvious difference is certainly the presence of interparapodial lateral pouches in A. insolita View in CoL comb. nov. from chaetiger 6, this character is missing in its congener. Comparison of specimen length in relation to the number of chaetigers shows that specimens of A. insolita View in CoL comb. nov. are longer on average than specimens of A. dayi View in CoL from the Atlantic seamounts with the same number of chaetigers.
Size-dependent characters were found in both species (Figure 5). The larger (determined by body width) the specimen the more branchial pairs are present. Branchial length (also in relation to the length of the notopodial postchaetal lamellae) is positively correlated with specimen size. Neuropodial hooks are first present in more posterior chaetigers in larger specimens than in smaller ones. The maximum number of hooks in both parapodial rami increases with specimen size. An examination of the pattern of dorsal holes could only be undertaken for very few specimens of A. insolita View in CoL comb. nov., but the general pattern is the same as described for A. dayi View in CoL from the Atlantic seamounts.
Another Aonidella View in CoL species ocurring in the Atlantic Ocean is A. cirrobranchiata ( Day, 1961) View in CoL . The species is easily distinguished from A. dayi View in CoL by the presence of bidentate hooks with one apical tooth only (opposed to at least two apical teeth in A. dayi View in CoL ). In the course of the present study we also examined a single specimen of A. cirrobranchiata View in CoL from the Bay of Biscay (USNM 80482). The specimen is an anterior fragment of about 20 chaetigers with only two hook-bearing chaetigers and, most importantly, exhibits interparapodial lateral pouches from between chaetigers 6/7 until the end of the fragment. The specimen was studied by Maciolek (2000) together with material from South Africa, but the presence of pouches was not mentioned in her description. López (2010) reported A. cirrobranchiata View in CoL from the Bellingshausen Sea observing only slight differences to the original description and previous reports in the newly collected material. The occurrence of pouches was neither observed in specimens from the Southern Ocean nor in those from South Africa ( López 2010). Except for the presence of interparapodial lateral pouches the specimen from the Bay of Biscay agrees well with former descriptions of A. cirrobranchiata View in CoL though it has to be considered that it is incomplete and not very well preserved. Interparapodial lateral pouches are present in Aonidella insolita View in CoL comb. nov. from Western Australia. A. insolita View in CoL comb. nov. is easily distinguished from A. cirrobranchiata View in CoL by the presence of hooks with two apical teeth instead of only one.
In general, all currently known Aonidella View in CoL spp. are of great morphological similarity. Except for the presence or absence of pouches and the number of apical teeth in neuropodial and notopodial hooded hooks there are no distinct feature for the different species. The study of size-dependent characters should be undertaken with care as well as the search for new characters.
FIGURE 5. Correlation between body width (measured at 10th chaetiger) and last branchiate chaetiger (left), or first appearance of neuropodial hooks (right), in Aonidella View in CoL spp. from both sides of the Atlantic Ocean and from the Indian Ocean (off Western Australia).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
InfraClass |
Canalipalpata |
|
Order |
|
|
Family |
|
|
Genus |
Aonidella cf. dayi Maciolek
| Bick, Andreas, Guggolz, Theresa & Götting, Miriam 2014 |
Prionospio (Minuspio) cf. cirrobranchiata
| Maciolek 2000: 531 |
| Imajima 1992: 57 |
| Lopez-Jamar 1989: 107 |
| Johnson 1984: 6 |
| Day 1973: 73 |