
Pugettia vulgaris, Ohtsuchi, Naoya, Kawamura, Tomohiko & Takeda, Masatsune, 2014
|
publication ID |
https://doi.org/10.11646/zootaxa.3765.6.4 |
|
publication LSID |
lsid:zoobank.org:pub:3E9B7388-CF58-46D2-8A37-E820E165772D |
|
DOI |
https://doi.org/10.5281/zenodo.6134577 |
|
persistent identifier |
https://treatment.plazi.org/id/03D6E818-FFB8-6A13-85E4-FE5BFCEAF9EB |
|
treatment provided by |
Plazi |
|
scientific name |
Pugettia vulgaris |
| status |
sp. nov. |
Pugettia vulgaris View in CoL n. sp.
[New Japanese name: Arasaki-mo-gani] ( Figs. 4 View FIGURE 4 , 5 View FIGURE 5 , 6 View FIGURE 6 F–J, 7)
Pugettia quadridens pellucens: Marumura & Kosaka 2003: 32 View in CoL (in part).
Material examined. Holotype: NSMT-Cr 22172, male (CW 5.8 mm, PCL 7.8 mm), Nagai, Yokosuka, Miura Peninsula, 7–8 m, articulated coralline turfs on rocky reefs, 18 May 2010, SCUBA diving, coll. J. Hayakawa & N. Ohtsuchi.
Allotype: NSMT-Cr 22173, ovigerous female (CW 5.3 mm, PCL 6.8 mm), same data as holotype.
Paratypes: NSMT-Cr 22174, 2 males (CW 2.7, 4.2 mm, PCL 3.8, 6.0 mm), 1 female (CW 3.5 mm, PCL 5.2 mm), same data as holotype.—NSMT-Cr 22175, 1 male (CW 7.3 mm, PCL 9.9 mm), 1 female (CW 3.3 mm, PCL 5.1 mm), 1 ovigerous female (CW 6.4 mm, PCL 8.2 mm), same locality, 2–4 m, articulated coralline turfs on rocky reef, 24 April 2009, SCUBA diving, coll. J. Hayakawa & N. Ohtsuchi.—CBM-ZC 11019, 2 females (CW 3.4, 6.9 mm, PCL 4.7, 9.2 mm), same locality, 2–4 m, gelidiacean turfs on rocky reef, 24 April 2009, SCUBA diving, coll. J. Hayakawa & N. Ohtsuchi.—CBM-ZC 11020, 1 male (CW 4.1 mm, PCL 6.1 mm), 1 ovigerous female (CW 6.0 mm, PCL 7.0 mm), 2 females (CW 3.7, 5.3 mm, PCL 5.0, 5.9 mm), same locality, 2–4 m, gelidiacean turfs on rocky reefs, 17 June 2009, SCUBA diving, coll. J. Hayakawa & N. Ohtsuchi.
Non-type: CBM-ZC 11022, 1 male (CW 9.8 mm, PCL 13.0 mm), 1 female (CW 10.9 mm, PCL 13.4 mm) Tomiyama Fishery Port, raft for aquaculture, 29 December 1995, hand, coll. T. Komai.—WPNH-Na-Cr 0313-3, 1 male (CW 5.1 mm, PCL 8.0 mm), off Shionomisaki, Wakayama, coll. S. Nagai (examined by Marumura & Kosaka 2003).
The following specimens were all collected from Nagai, Yokosuka, Miura Peninsula, using SCUBA diving by J. Hayakawa & N. Ohtsuchi and deposited in AORI; 3 males (CW 2.1–4.8 mm, PCL 3.2–6.6 mm), 2 females (CW 2.4, 2.6 mm, PCL 3.8, 4.3 mm), 2–4 m, gelidiacean turfs on rocky reef, 10 December 2008.— 4 males (CW 2.1–5.3 mm, PCL 3.5–7.4 mm), 2 females (CW 3.6, 4.6 mm, PCL 5.2, 6.5 mm), the same habitat, 27 January 2009.— 1 male (CW 3.0 mm, PCL 4.1 mm), 2–4 m, articulated coralline algal turfs on rocky reef, 11 September 2009.— 2 males (CW 2.4, 5.5 mm, PCL 3.2, 7.0 mm), 1 female (CW 5.2 mm, PCL 6.7 mm), 7–8 m, articulated coralline algal turfs on rocky reef, 15 March 2009.— 1 female (CW 6.0 mm, PCL 8.5 mm), 2 ovigerous females (CW 5.4, 5.8 mm, PCL 7.3, 8.1 mm), the same habitat, 25 May 2009.— 3 males (CW 5.0– 6.7 mm, PCL 7.3–9.2 mm), 2 females (CW 4.9, 5.8 mm, PCL 7.6, 7.8 mm),1 ovigerous females (CW 4.8 mm, PCL 6.4 mm), the same habitat, 17 June 2009.— 2 males (CW 2.6, 5.9 mm, PCL 3.3, 8.2 mm), 1 female (CW 5.0 mm, PCL 6.9 mm), the same habitat, 5 April 2010.— 3 males (CW 4.4–6.0 mm, PCL 6.0– 8.1 mm), 2 females (CW 4.3, 4.4 mm, PCL 6.4, 6.1 mm), the same habitat, 26 April 2010.
Etymology. The name is derived from the Latin vulgaris for “common ”, alluding to its high capture frequency in the type locality.
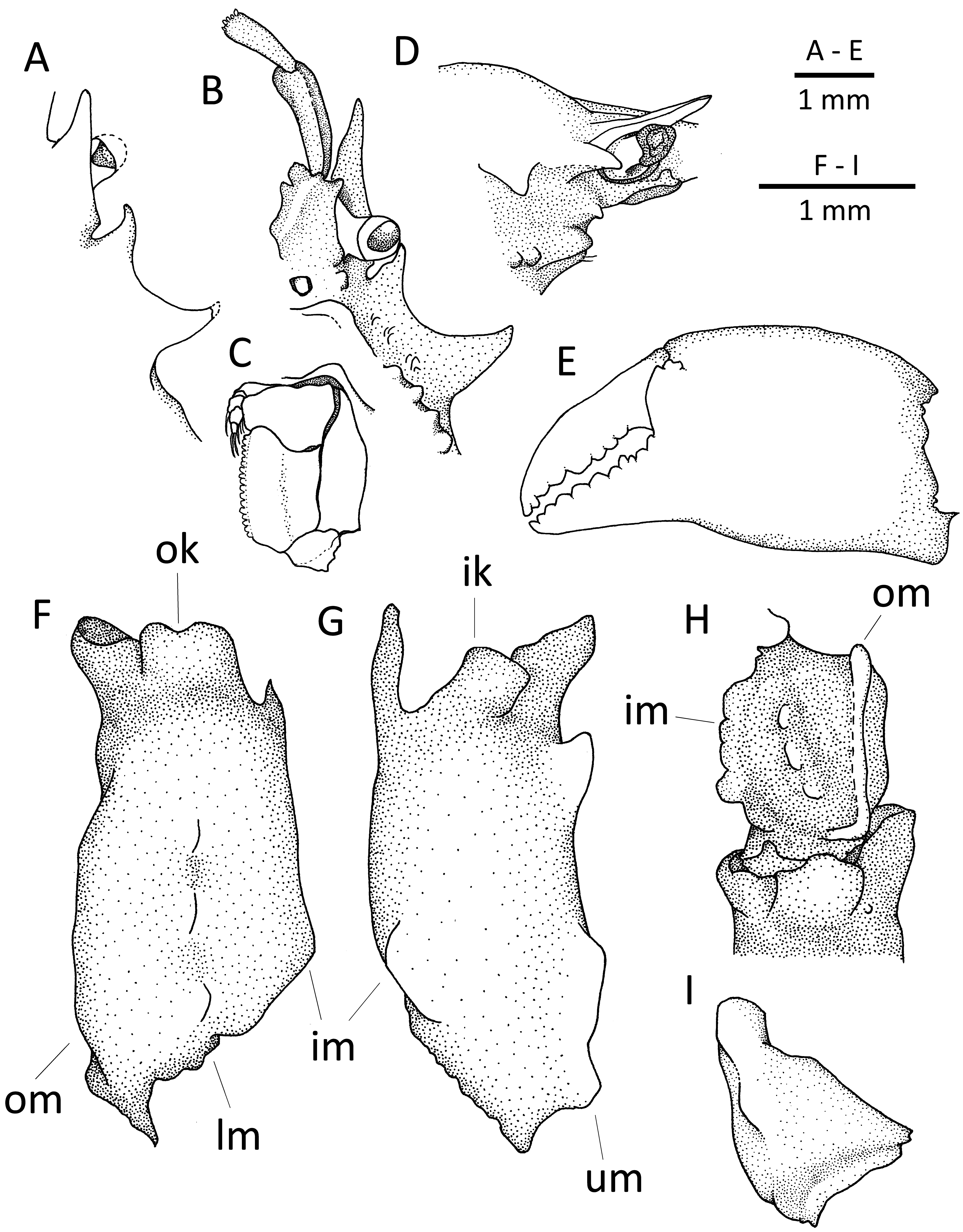
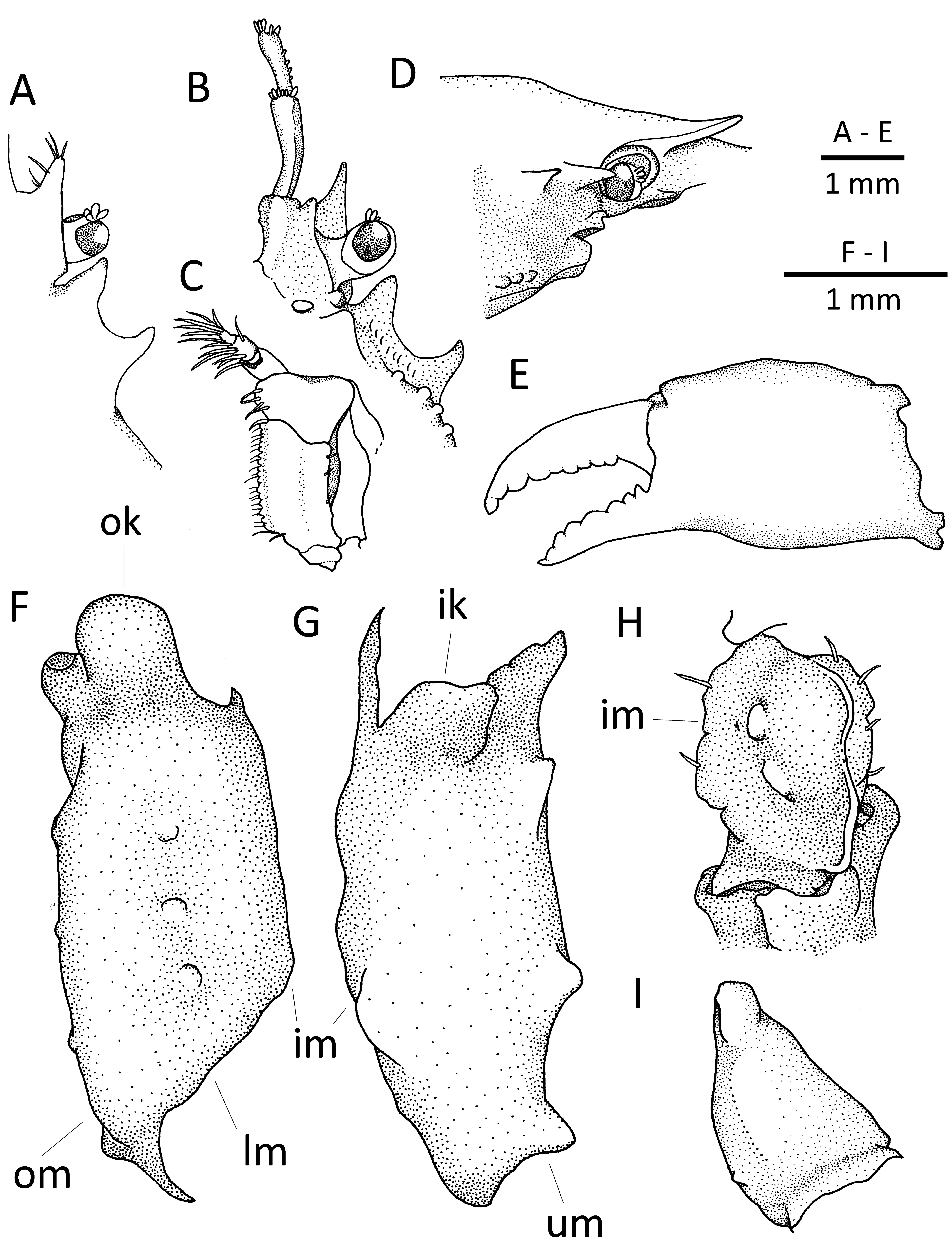
Description. Carapace ( Figs. 4 View FIGURE 4 A, 7) elongated pyriform; regions not markedly defined; gastric, cardiac, branchial regions moderately elevated; protogastric region with 2 oblique rows of hooked setae on either side of midline; intestinal region weakly elevated. Tufts of a few elongated setae present on midline of protogastric region midpoint of epibranchial region, apex of epibranchial spine, summits of cardiac region and intestinal region ( Fig. 4 View FIGURE 4 A); dorsal surface sometimes entirely covered with dense club-shaped setae; no conspicuous tubercles or spines on midline of gastric, cardiac, intestinal regions.
Rostral spines ( Figs. 4 View FIGURE 4 A, 7) moderately long, about 0.3 of postrostral carapace length (0.34±0.04, N = 12), moderately diverging anteriorly, each with 2 rows of hooked setae in proximal 0.5–0.7. Preorbital spine ( Figs. 4 View FIGURE 4 A, 5A, B, D, 7) moderately strong, slightly diverging anteriorly; lateral margin of supraorbital eave straight. Orbital hiatus ( Figs. 4 View FIGURE 4 A, 5A) deep, distinctly separating supraorbital eave from postorbital lobe. Postorbital lobe ( Figs. 4 View FIGURE 4 , 5 View FIGURE 5 A, B, D) compressed dorsoventrally, triangular with acute apex, directed anterolaterally in dorsal view, horizontal in lateral view. Hepatic lobe ( Figs. 4 View FIGURE 4 , 5 View FIGURE 5 A, B, D) compressed, acuminate, subequal in size to slightly smaller than postorbital lobe, slightly fused at base with postorbital lobe, projecting anterolaterally, distally curved anteriorly. Concavity between postorbital and hepatic lobes U-shaped ( Figs. 4 View FIGURE 4 , 5 View FIGURE 5 A, B). Anterolateral margin of carapace ( Figs. 4 View FIGURE 4 A, 7) with rows of hooked setae; lateral surface inferior to anterolateral margin with two small tubercles. Epibranchial spines ( Figs. 4 View FIGURE 4 A, 7) acute, located at posterior 0.3 of postrostral carapace length.
Subhepatic region ( Figs. 4 View FIGURE 4 B, 5B) unarmed. Pterygostomial region ( Figs. 4 View FIGURE 4 B, 5B) not particularly inflated, with 4 tubercles along pleural suture.
Basal antennal article ( Fig. 5 View FIGURE 5 B) with ventral surface unarmed, bearing low, blunt longitudinal ridge mesial to mi line; distolateral angle moderately produced into small subacute spine directed anterolaterally; lateral margin faintly sinuous, with small tubercle basally. Antennal peduncle consisting of 2 articles flattened dorsoventrally; penultimate article ( Figs. 4 View FIGURE 4 B, 5B) with lateral margin carinated over entire length; ultimate article distinctly shorter than penultimate article, proximal end almost as broad as distal end.
Third maxilliped ( Fig. 5 View FIGURE 5 C) unarmed on surface; ischium with shallow median groove; anterolateral angle of merus produced, slightly upturned; exopod unarmed.
Anterolateral angle of buccal frame ( Figs. 5 View FIGURE 5 C, D) moderately produced, overlapped by anterolateral angle of merus of third maxilliped when closed.
Chelipeds ( Figs. 4 View FIGURE 4 , 5 View FIGURE 5 E–I, 7) equal in size, similar in shape. Ischium ( Figs. 4 View FIGURE 4 B, 5I) weakly swollen ventrally in distal half; mesial margin slightly ridged; distolateral lobe distinct. Merus ( Figs. 4 View FIGURE 4 , 5 View FIGURE 5 F, G) prismatic; dorsal surface ( Figs. 4 View FIGURE 4 A, 5G) bearing 3 lamellar teeth or lobes in proximal 0.8; outer surface ( Figs. 4 View FIGURE 4 A, 5F) with obtuse longitudinal ridge with 3 low tubercles; ventral surface ( Figs. 4 View FIGURE 4 B, 5F) with bluntly ridge bearing 3 low tubercles; upper inner surface ( Figs. 4 View FIGURE 4 , 5 View FIGURE 5 G) generally unarmed, bluntly ridged; inner margin ( Figs. 4 View FIGURE 4 , 5 View FIGURE 5 G) armed with low convexity proximally; distal margin with 2 prominent knobs at articulation with carpus (outer knob larger than inner); prominent, obliquely erect, rounded or subrectangular lobe on upper side ( Figs. 4 View FIGURE 4 A, 5F, G). Carpus ( Fig. 4 View FIGURE 4 A, 5H) weakly inflated with distinct, bi- or tri-tuberculate ridge on upper surface; inner margin crested, divided into 2 faint lobes; outer margin distinctly crested. Chela ( Fig. 5 View FIGURE 5 E) almost twice longer than high (length/ height = 2.0–2.3, N = 4) in adult males; upper margin weakly crested, with small rounded lobe basally; lower margin slightly ridged.
Ambulatory legs ( Figs. 4 View FIGURE 4 , 7) decreasing in length posteriorly. Meri each with small upper distal tubercle, otherwise unarmed. Carpi each with shallow depression on upper surface. Dactyli each with 2 rows of small calcareous spines on flexor surface.
Male first gonopod ( Fig. 6 View FIGURE 6 F–J) similar to that of P. pellucens , but dorsal lobe not elongate, crossing mesial lobe even in large specimens; hiatus between dorsal and mesial lobe always narrow.
Variation. Hepatic region of carapace relatively more strongly elevated in females (Fig. 7A) than in males. Anterior margin of postorbital lobe with a few spinules in small specimens (<CW 2.7 mm). Hepatic lobe platelike, similar to postorbital lobe in size and shape in small specimens (<CW 4.2 mm, Fig. 7B). Number of lobes on inner margin of the cheliped carpus two or three in adults, but absent in small specimens (<CW 2.7 mm). In large males (> CW 5.5 mm), chela relatively larger compared to small males and females ( Figs. 4 View FIGURE 4 , 7). First gonopod of male specimens larger than CW 4.1 mm similar to that of holotype ( Fig. 6 View FIGURE 6 ), while in small males (<CW 3.0 mm), shaft not folded, apex not trilobate.
Size. Largest male PCL 13.0 mm, CW 9.8 mm, largest female PCL 13.4 mm, CW 10.9 mm, smallest ovigerous female PCL 6.4 mm, CW 4.8 mm.
Camouflage. Composed of various combinations of sponges, hydrozoans, bryzoans, pieces of the red algae (Rhodophyta), which are abundant in the type locality.
Habitat. Occurring in shallow water from intertidal to 8 m deep. In Nagai, Sagami Bay, the crab was found to inhabit gelidiacean red algal turfs of Gelidium elegans and/or Ptelocladia spp., and articulated coralline algal turfs of Amphiroa dilatata , A. zonata , Corallina officinalis , C. pilulifera , and Marginisporum crassissima developed on the subtidal rocky reefs. The new species is the predominant brachyuran species in the red algal turfs in shallow subtidal areas of Nagai (N. Ohtsuchi, personal observation).
Distribution. At present known with certainty only from Japan, from Boso Peninsula to Kii Peninsula.
Remarks. Pugettia vulgaris n. sp. appears closest to P. pellucens in having a more elongated pyriform carapace, narrow compressed supraorbital eave with an acute preorbital spine, clearly separated postorbital and hepatic lobes, cheliped merus with a tridentate dorsal margin and carpus bearing crested inner and outer margins. Differentiating characters between the two species are discussed below.
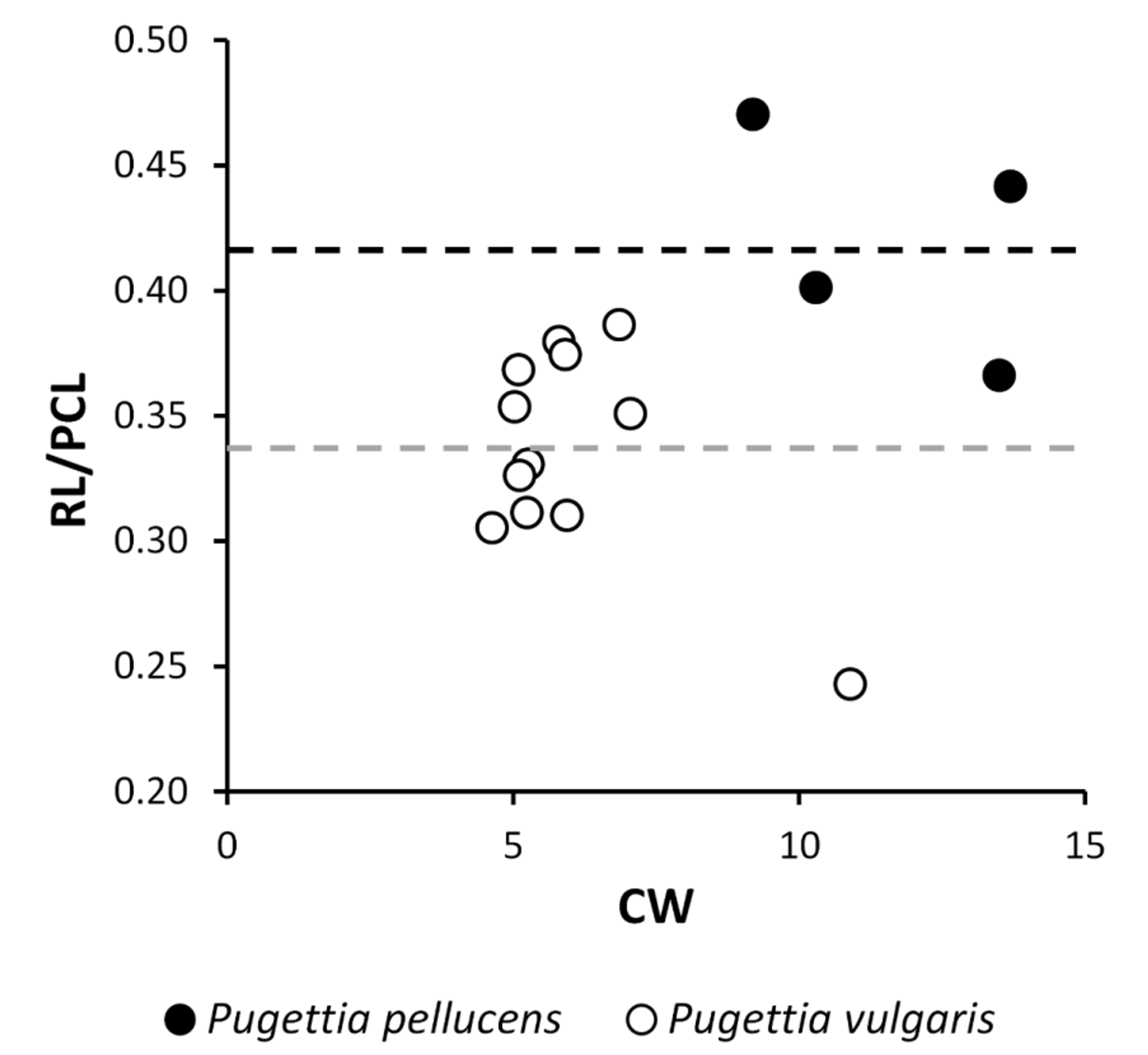
The rostral spines tend to be shorter in P. vulgaris n. sp. ( Figs. 4 View FIGURE 4 , 7) than in P. pellucens ( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 ), although the ranges of variation partially overlap between the two species. The relative length of the rostral spines against the postrostral carapace length (RL/PCL) ranges from 0.24–0.39 (N = 12) in P. vulgaris n. sp., but from 0.36–0.47 in P. pellucens (N = 4). The mean RL/PCL is significantly different between the two species ( Fig. 8 View FIGURE 8 , 0.34± 0.04 in P. vulgaris n. sp. versus 0.42± 0.04 in P. pellucens , two sample t -test, p <0.01).
The hepatic lobe is subequal in the size to or slightly larger than the postorbital lobe and projects anterolaterally in P. vulgaris n. sp. ( Fig. 4 View FIGURE 4 A) rather than being distinctly larger than the latter, and projecting laterally in P. pellucens ( Figs. 1 View FIGURE 1 A, 2B).
The penultimate segment of the antennal peduncle is relatively slender in P. vulgaris n. sp. ( Fig. 5 View FIGURE 5 B) than in P. pellucens ( Fig. 3 View FIGURE 3 B).
The buccal frame is moderately produced anterolaterally and the anterolateral angle is overlapped by the anterolateral angle of the merus of the third maxilliped (when closed) in P. vulgaris n. sp. ( Figs. 5 View FIGURE 5 C, D); whereas in P. pellucens , the buccal frame is strongly produced and the anterolateral angle is not overlapped by the anterolateral angle of the merus of third maxilliped when closed ( Figs. 3 View FIGURE 3 C, D).
The cheliped ischium is always rounded mesially in P. vulgaris n. sp. ( Figs. 4 View FIGURE 4 , 5 View FIGURE 5 I), whereas it can have a blunt ridge on the mesial surface in P. pellucens ( Figs. 1 View FIGURE 1 A, B, 2A, 3I). This character, however, should be used with caution because of its variability in the latter species.
Field studies at Nagai, Yokosuka, Miura Peninsula, by the senior author, have shown that Pugettia vulgaris n. sp. is restricted to red algal turfs on the rocky reef in intertidal to subtidal depth of 8 m. On the other hand, the present specimens of P. pellucens were collected from mud, gravel or rock bottoms at depths of 5– 60 m. No specimens referable to P. pellucens were found in algal habitats where P. vulgaris were abundant so the two taxa do not appear to be sympatric. The possible difference in the habitat preference supports the recognition of P. vulgaris n. sp. as a distinct species from P. pellucens .
Pugettia vulgaris View in CoL n. sp. is also morphologically similar to P. elongata Yokoya, 1933 View in CoL , and P. intermedia Sakai, 1938 View in CoL , particularly in having partly fused, subequal postorbital and hepatic lobes (cf. Griffin & Tranter 1986). Small immature specimens of P. vulgaris View in CoL n. sp. (CW <5.0 mm) may be confused with these species.
Pugettia elongata View in CoL is so far represented only by two male syntypes from Omae-Zaki Light, Suruga Bay, Japan ( Yokoya 1933) and four males from off Choshi and Shionomisaki, Japan ( Marumura & Kosaka 2003: 32). Unfortunately, our efforts to locate these specimens were not successful. Comparison with the original description of P. elongata View in CoL by Yokoya (1933), nevertheless, revealed that P. elongata View in CoL is distinguished from P. vulgaris View in CoL n. sp. as follows: P. elongata View in CoL has a spiniform median tubercle on the gastric, cardiac and intestinal regions of the carapace, respectively (cf. Yokoya 1933: text-fig. 54), whereas in P. vulgaris View in CoL n. sp. such tubercles are absent on the carapace ( Figs. 4 View FIGURE 4 A, 7); the concavity separating the postorbital and hepatic lobes are very shallow in P. elongata View in CoL (cf. Yokoya 1933: text-fig. 54), but the concavity is deep in P. vulgaris View in CoL n. sp. ( Figs. 4 View FIGURE 4 A, 5A); the upper margin of the cheliped merus is lined with an irregularly dentate carina in P. elongata View in CoL (cf. Yokoya 1933: text-fig. 54), whereas in P. vulgaris View in CoL n. sp. the margin is not carinate, but armed with three lamellar teeth or lobes ( Figs. 4 View FIGURE 4 A, 5G).
Pugettia intermedia View in CoL can be distinguished from P. vulgaris View in CoL n. sp. by the following characters. In P. intermedia View in CoL , carapace regions are well-defined, of them the gastric, cardiac and mesobranchial regions are elevated, armed with four, two, and one tubercles, respectively ( Sakai 1938: pl. 36, fig. 2). In P. vulgaris View in CoL n. sp., these regions are not markedly elevated, and without armature ( Fig. 4 View FIGURE 4 A). The epibranchial spines are also proportionately stronger, projecting backwards and upwards in P. intermedia View in CoL ( Sakai 1938: pl. 36, fig. 2), whereas in P. vulgaris View in CoL n. sp., they are distinctly weaker and projecting laterally ( Figs. 4 View FIGURE 4 A, 7). The cheliped merus has a longitudinal ridge bearing five teeth on the outer surface in P. intermedia View in CoL ( Sakai 1938: pl. 36, fig. 2), but there are only three low tubercles in P. vulgaris View in CoL n. sp. ( Fig. 5 View FIGURE 5 F). The tip of mesial lobe of the male first gonopod points distinctly lower than that of the dorsal lobe in P. intermedia View in CoL (cf. Griffin & Tranter 1986: 94, fig. 28a, b; Ariyama 1995: 3, fig. 3a–c), whereas in P. vulgaris View in CoL n. sp., the tip exceeds that of the dorsal lobe ( Fig. 6 View FIGURE 6 F–J).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Brachyura |
|
SuperFamily |
Majoidea |
|
Family |
|
|
SubFamily |
Epialtinae |
|
Genus |
Pugettia vulgaris
| Ohtsuchi, Naoya, Kawamura, Tomohiko & Takeda, Masatsune 2014 |
Pugettia quadridens pellucens :
| Marumura 2003: 32 |